

Control of intracellular heme levels: heme transporters and heme oxygenases
- PMID: 21238504
- PMCID: PMC3079059
- DOI: 10.1016/j.bbamcr.2011.01.008
Control of intracellular heme levels: heme transporters and heme oxygenases
Abstract
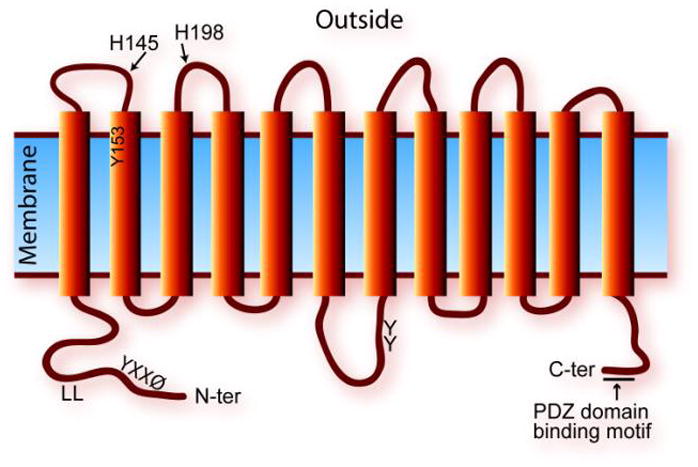
Heme serves as a co-factor in proteins involved in fundamental biological processes including oxidative metabolism, oxygen storage and transport, signal transduction and drug metabolism. In addition, heme is important for systemic iron homeostasis in mammals. Heme has important regulatory roles in cell biology, yet excessive levels of intracellular heme are toxic; thus, mechanisms have evolved to control the acquisition, synthesis, catabolism and expulsion of cellular heme. Recently, a number of transporters of heme and heme synthesis intermediates have been described. Here we review aspects of heme metabolism and discuss our current understanding of heme transporters, with emphasis on the function of the cell-surface heme exporter, FLVCR. Knockdown of Flvcr in mice leads to both defective erythropoiesis and disturbed systemic iron homeostasis, underscoring the critical role of heme transporters in mammalian physiology. This article is part of a Special Issue entitled: 11th European Symposium on Calcium.
2011 Elsevier B.V. All rights reserved.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
