

Towards a more nuanced understanding of the relationship between sex-biased gene expression and rates of protein-coding sequence evolution
- PMID: 21239389
- PMCID: PMC3098513
- DOI: 10.1093/molbev/msr010
Towards a more nuanced understanding of the relationship between sex-biased gene expression and rates of protein-coding sequence evolution
Abstract
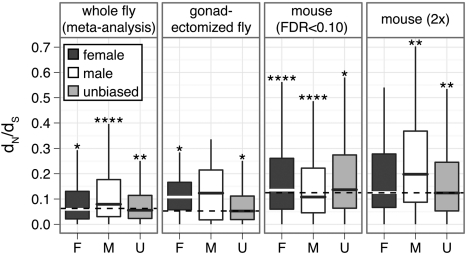
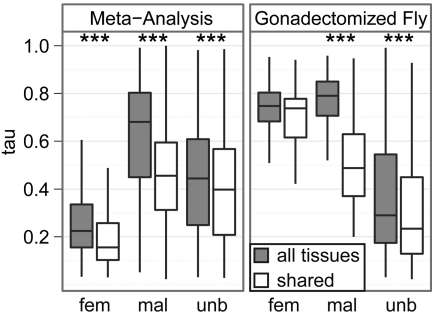
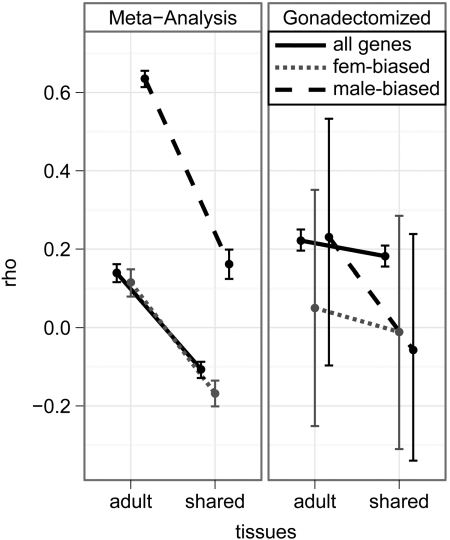
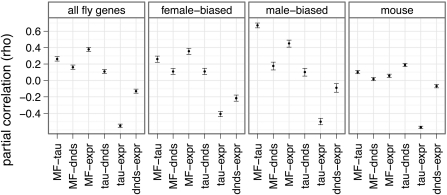
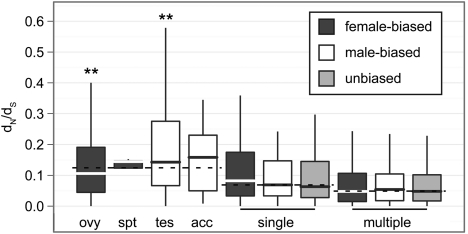
Genes that are differentially expressed between the sexes (sex-biased genes) are among the fastest evolving genes in animal genomes. The majority of sex-biased expression is attributable to genes that are primarily expressed in sex-limited reproductive tissues, and these reproductive genes are often rapidly evolving because of intra- and intersexual selection pressures. Additionally, studies of multiple taxa have revealed that genes with sex-biased expression are also expressed in a limited number of tissues. This is worth noting because narrowly expressed genes are known to evolve faster than broadly expressed genes. Therefore, it is not clear whether sex-biased genes are rapidly evolving because they have sexually dimorphic expression, because they are expressed in sex-limited reproductive tissues, or because they are narrowly expressed. To determine the extend to which other confounding variables can explain the rapid evolution of sex-biased genes, I analyzed the rates of evolution of sex-biased genes in Drosophila melanogaster and Mus musculus in light of tissue-specific measures of expression. I find that genes with sex-biased expression in somatic tissues shared by both sexes are often evolving faster than non-sex-biased genes, but this is best explained by the narrow expression profiles of sex-biased genes. Sex-biased genes in sex-limited tissues in D. melanogaster, however, evolve faster than other narrowly expressed genes. Therefore, the rapid evolution of sex-biased genes is limited only to those genes primarily expressed in sex-limited reproductive tissues.
Figures
References
-
- Arbeitman MN, Fleming AA, Siegal ML, Null BH, Baker BS. A genomic analysis of Drosophila somatic sexual differentiation and its regulation. Development. 2004;131:2007–2021. - PubMed
-
- Arbeitman MN, Furlong EEM, Imam F, Johnson E, Null BH, Baker BS, Krasnow MA, Scott MP, Davis RW, White KP. Gene expression during the life cycle of Drosophila melanogaster. Science. 2002;297:2270–2275. - PubMed
-
- Begun DJ, Lindfors HA. Rapid evolution of genomic Acp complement in the melanogaster subgroup of. Drosophila. Mol Biol Evol. 2005;22:2010–2021. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
