

Differential and complementary effects of glucose and prolactin on islet DNA synthesis and gene expression
- PMID: 21239441
- PMCID: PMC3198965
- DOI: 10.1210/en.2010-1258
Differential and complementary effects of glucose and prolactin on islet DNA synthesis and gene expression
Abstract
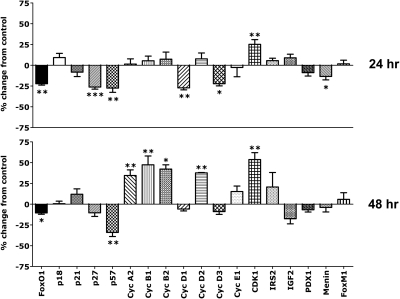
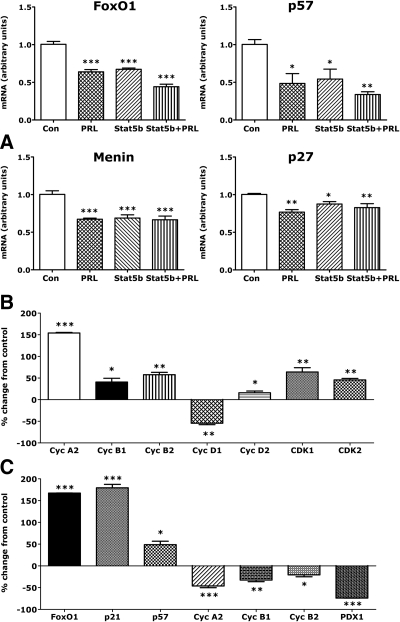
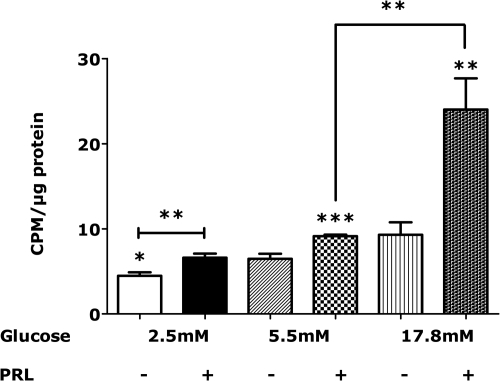
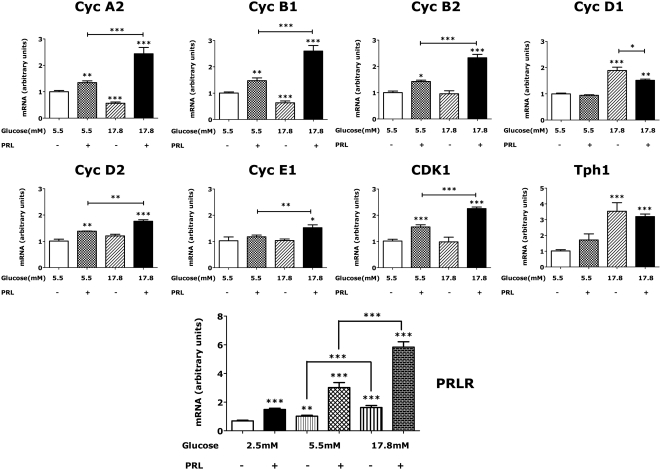
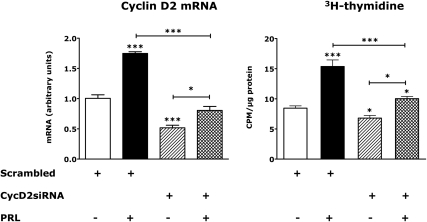
The mechanisms by which lactogenic hormones promote β-cell expansion remain poorly understood. Because prolactin (PRL) up-regulates β-cell glucose transporter 2, glucokinase, and pyruvate dehydrogenase activities, we reasoned that glucose availability might mediate or modulate the effects of PRL on β-cell mass. Here, we used male rat islets to show that PRL and glucose have differential but complementary effects on the expression of cell cyclins, cell cycle inhibitors, and various other genes known to regulate β-cell replication, including insulin receptor substrate 2, IGF-II, menin, forkhead box protein M1, tryptophan hydroxylase 1, and the PRL receptor. Differential effects on gene expression are associated with synergistic effects of glucose and PRL on islet DNA synthesis. The effects of PRL on gene expression are mirrored by β-cell overexpression of signal transducer and activator of transcription 5b and are opposed by dexamethasone. An ad-small interfering RNA specific for cyclin D2 attenuates markedly the effects of PRL on islet DNA synthesis. Our studies suggest a new paradigm for the control of β-cell mass and insulin production by hormones and nutrients. PRL up-regulates β-cell glucose uptake and utilization, whereas glucose increases islet PRL receptor expression and potentiates the effects of PRL on cell cycle gene expression and DNA synthesis. These findings suggest novel targets for prevention of neonatal glucose intolerance and gestational diabetes and may provide new insight into the pathogenesis of β-cell hyperplasia in obese subjects with insulin resistance.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
