

Malate plays a crucial role in starch metabolism, ripening, and soluble solid content of tomato fruit and affects postharvest softening
- PMID: 21239646
- PMCID: PMC3051241
- DOI: 10.1105/tpc.109.072231
Malate plays a crucial role in starch metabolism, ripening, and soluble solid content of tomato fruit and affects postharvest softening
Abstract
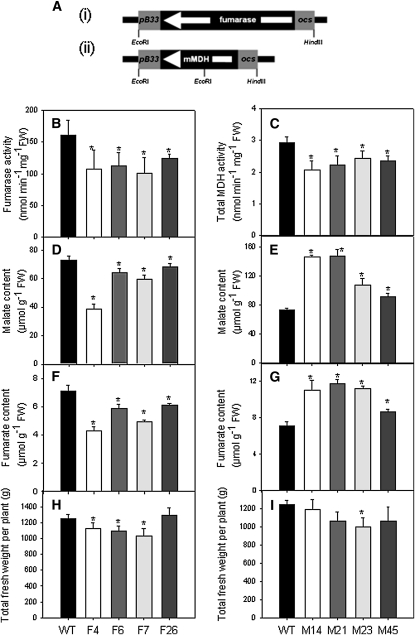
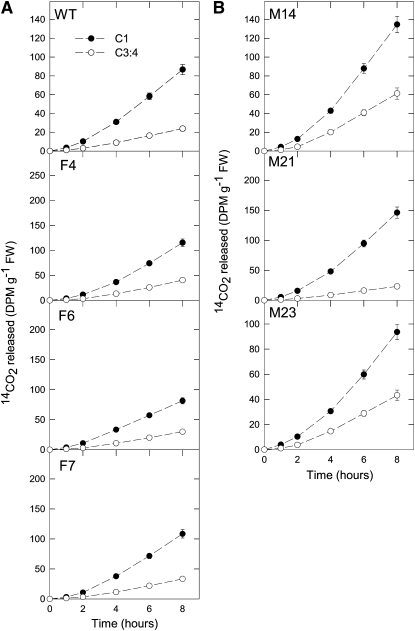
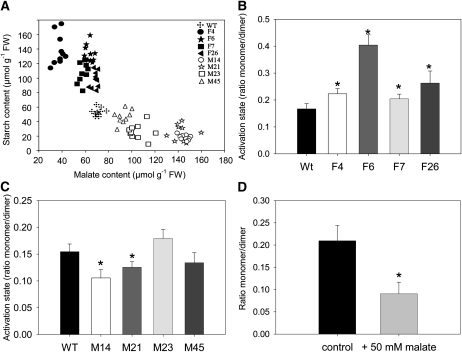
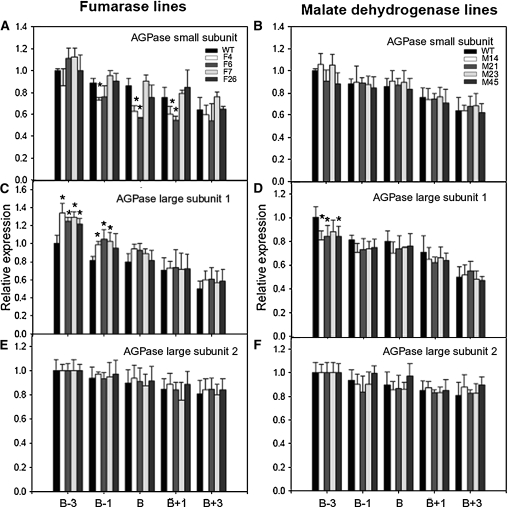
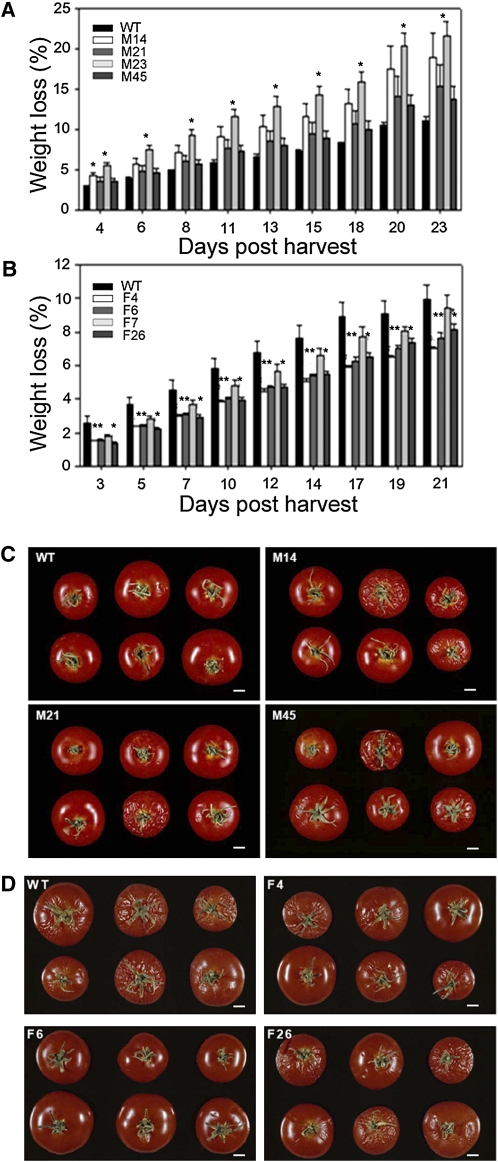
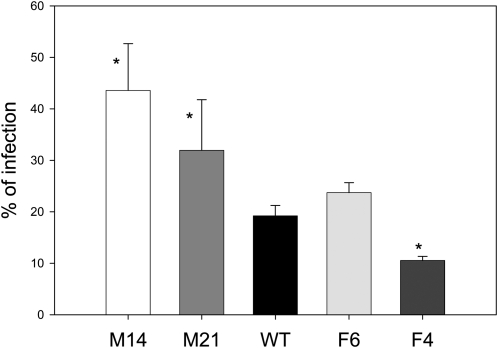
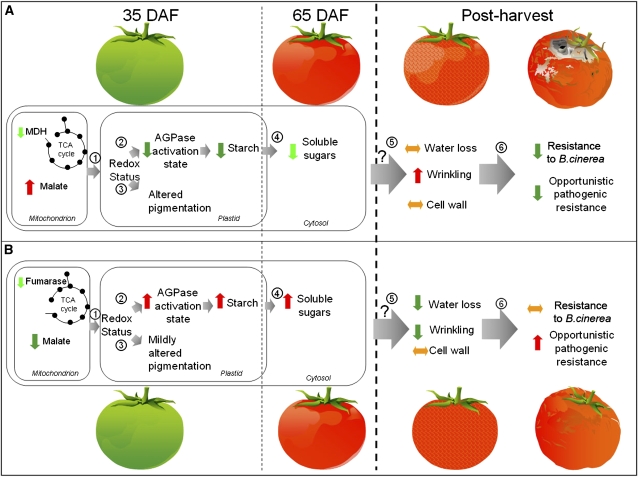
Despite the fact that the organic acid content of a fruit is regarded as one of its most commercially important quality traits when assessed by the consumer, relatively little is known concerning the physiological importance of organic acid metabolism for the fruit itself. Here, we evaluate the effect of modifying malate metabolism in a fruit-specific manner, by reduction of the activities of either mitochondrial malate dehydrogenase or fumarase, via targeted antisense approaches in tomato (Solanum lycopersicum). While these genetic perturbations had relatively little effect on the total fruit yield, they had dramatic consequences for fruit metabolism, as well as unanticipated changes in postharvest shelf life and susceptibility to bacterial infection. Detailed characterization suggested that the rate of ripening was essentially unaltered but that lines containing higher malate were characterized by lower levels of transitory starch and a lower soluble sugars content at harvest, whereas those with lower malate contained higher levels of these carbohydrates. Analysis of the activation state of ADP-glucose pyrophosphorylase revealed that it correlated with the accumulation of transitory starch. Taken together with the altered activation state of the plastidial malate dehydrogenase and the modified pigment biosynthesis of the transgenic lines, these results suggest that the phenotypes are due to an altered cellular redox status. The combined data reveal the importance of malate metabolism in tomato fruit metabolism and development and confirm the importance of transitory starch in the determination of agronomic yield in this species.
Figures
References
-
- Alba R., et al. (2004). ESTs, cDNA microarrays, and gene expression profiling: Tools for dissecting plant physiology and development. Plant J. 39: 697–714 - PubMed
-
- ap Rees T., Hill S.A. (1994). Metabolic control analysis of plant metabolism. Plant Cell Environ. 17: 587–599
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
