

HIF-1 modulates longevity and healthspan in a temperature-dependent manner
- PMID: 21241450
- PMCID: PMC3980873
- DOI: 10.1111/j.1474-9726.2011.00672.x
HIF-1 modulates longevity and healthspan in a temperature-dependent manner
Abstract
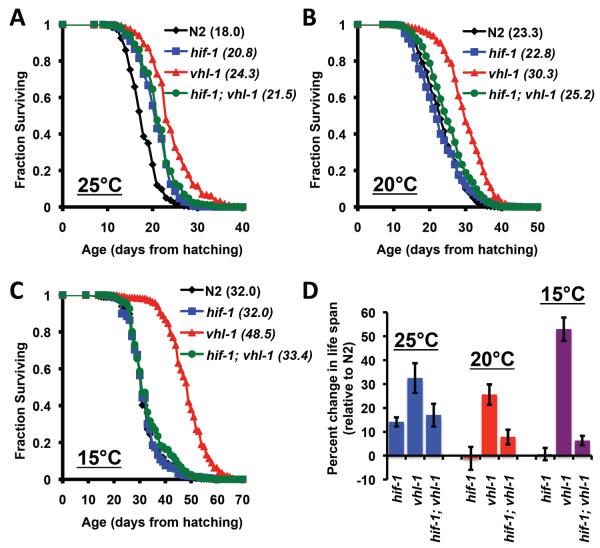
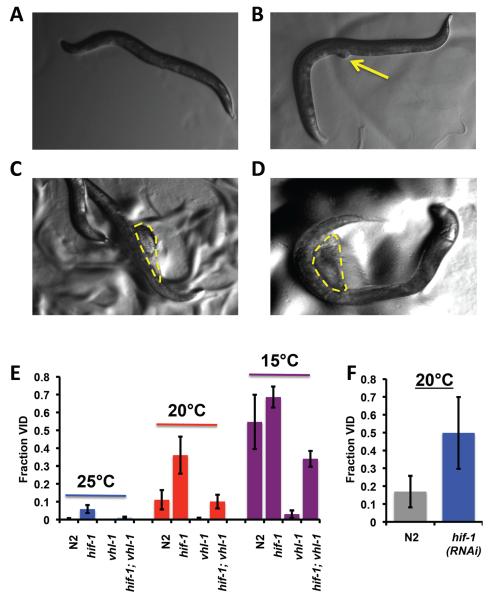
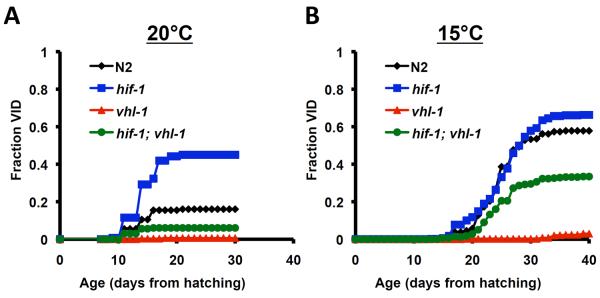
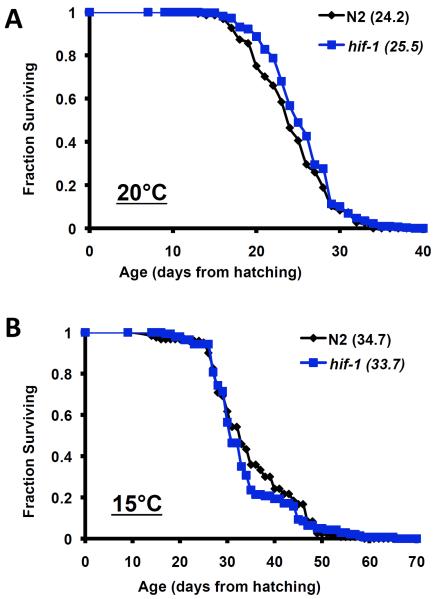
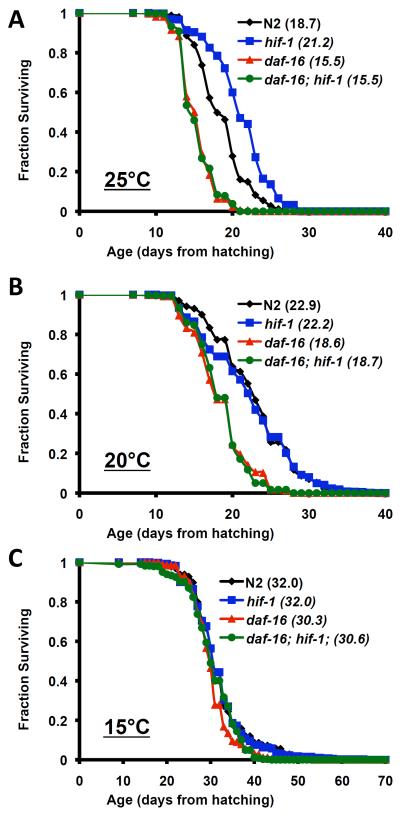
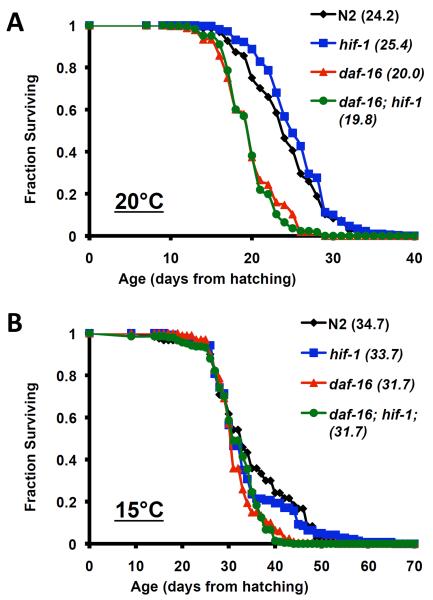
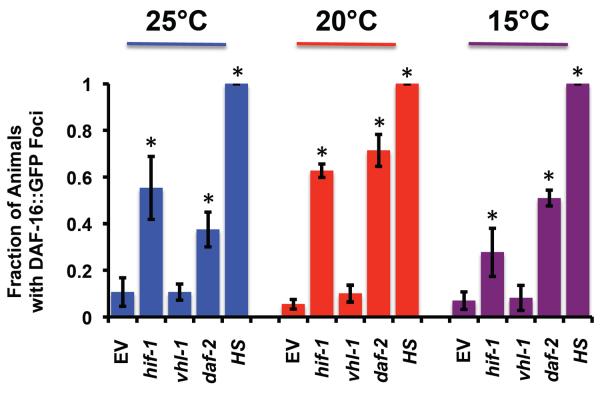
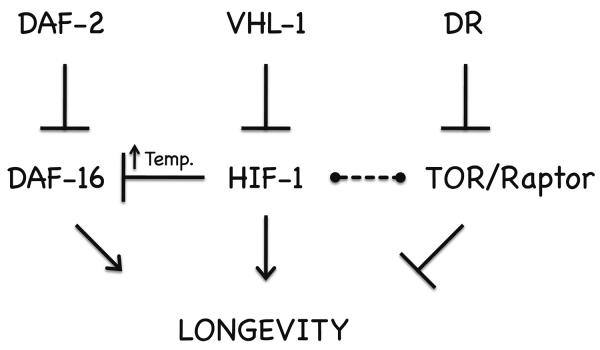
The hypoxia-inducible factor HIF-1 has recently been identified as an important modifier of longevity in the roundworm Caenorhabditis elegans. Studies have reported that HIF-1 can function as both a positive and negative regulator of life span, and several disparate models have been proposed for the role of HIF in aging. Here, we resolve many of the apparent discrepancies between these studies. We find that stabilization of HIF-1 increases life span robustly under all conditions tested; however, deletion of hif-1 increases life span in a temperature-dependent manner. Animals lacking HIF-1 are long lived at 25°C but not at 15°C. We further report that deletion or RNAi knockdown of hif-1 impairs healthspan at lower temperatures because of an age-dependent loss of vulval integrity. Deletion of hif-1 extends life span modestly at 20°C when animals displaying the vulval integrity defect are censored from the experimental data, but fails to extend life span if these animals are included. Knockdown of hif-1 results in nuclear relocalization of the FOXO transcription factor DAF-16, and DAF-16 is required for life span extension from deletion of hif-1 at all temperatures regardless of censoring.
© 2011 The Authors. Aging Cell © 2011 Blackwell Publishing Ltd/Anatomical Society of Great Britain and Ireland.
Figures
References
-
- Ahmetov II, Hakimullina AM, Lyubaeva EV, Vinogradova OL, Rogozkin VA. Effect of HIF1A gene polymorphism on human muscle performance. Bull Exp Biol Med. 2008;146:351–353. - PubMed
-
- Berdichevsky A, Viswanathan M, Horvitz HR, Guarente L. C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell. 2006;125:1165–1177. - PubMed
-
- Calnan DR, Brunet A. The FoxO code. Oncogene. 2008;27:2276–2288. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
