

RalB and the exocyst mediate the cellular starvation response by direct activation of autophagosome assembly
- PMID: 21241894
- PMCID: PMC3038590
- DOI: 10.1016/j.cell.2010.12.018
RalB and the exocyst mediate the cellular starvation response by direct activation of autophagosome assembly
Abstract
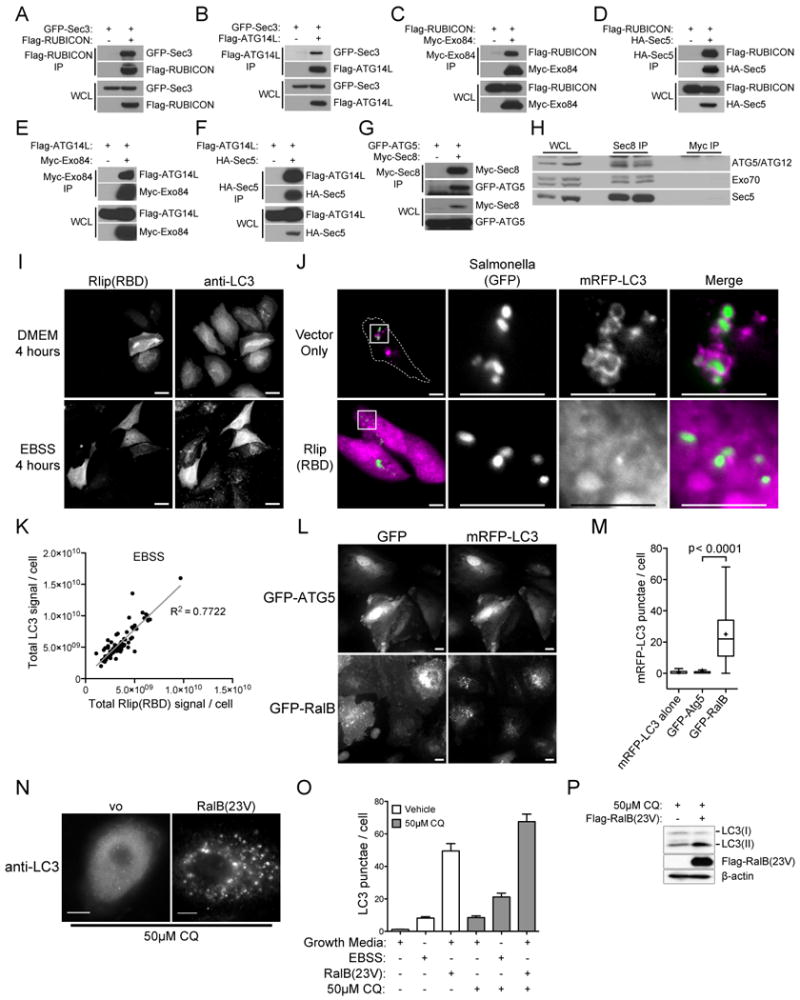
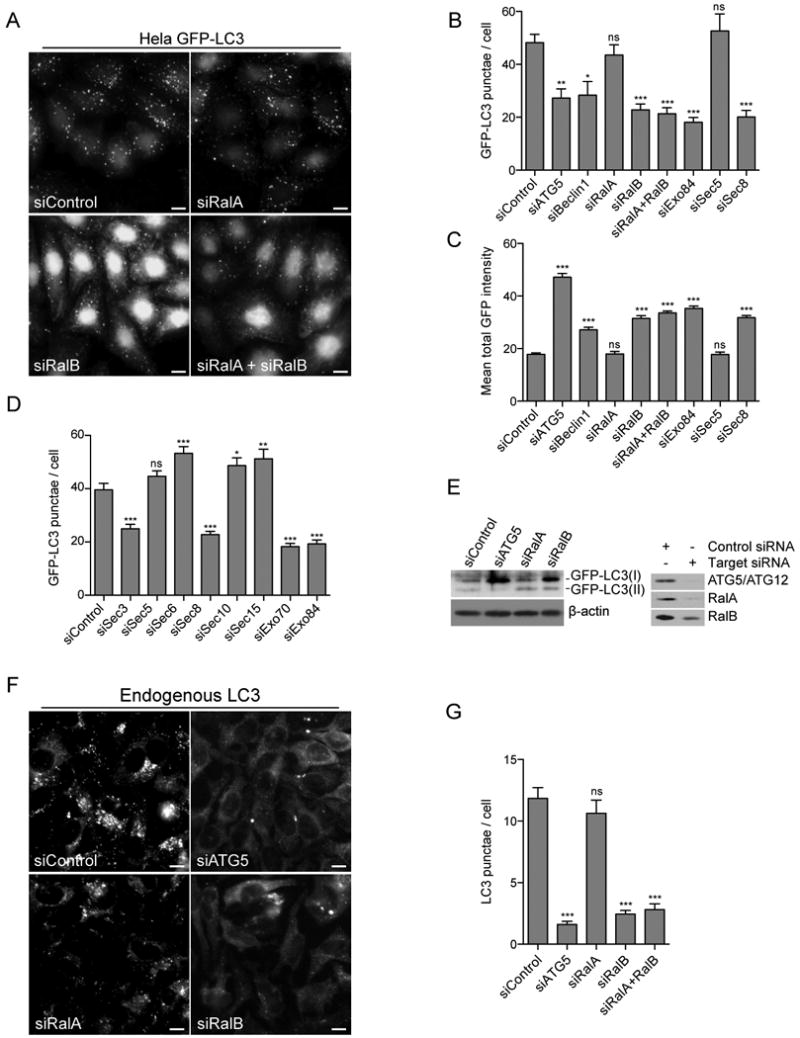
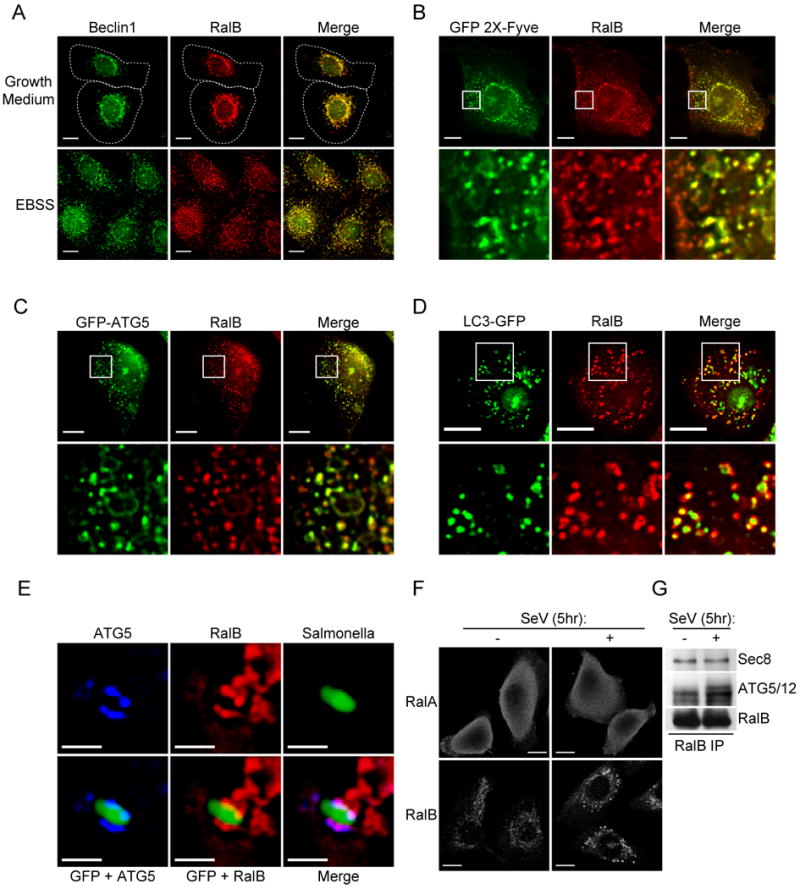
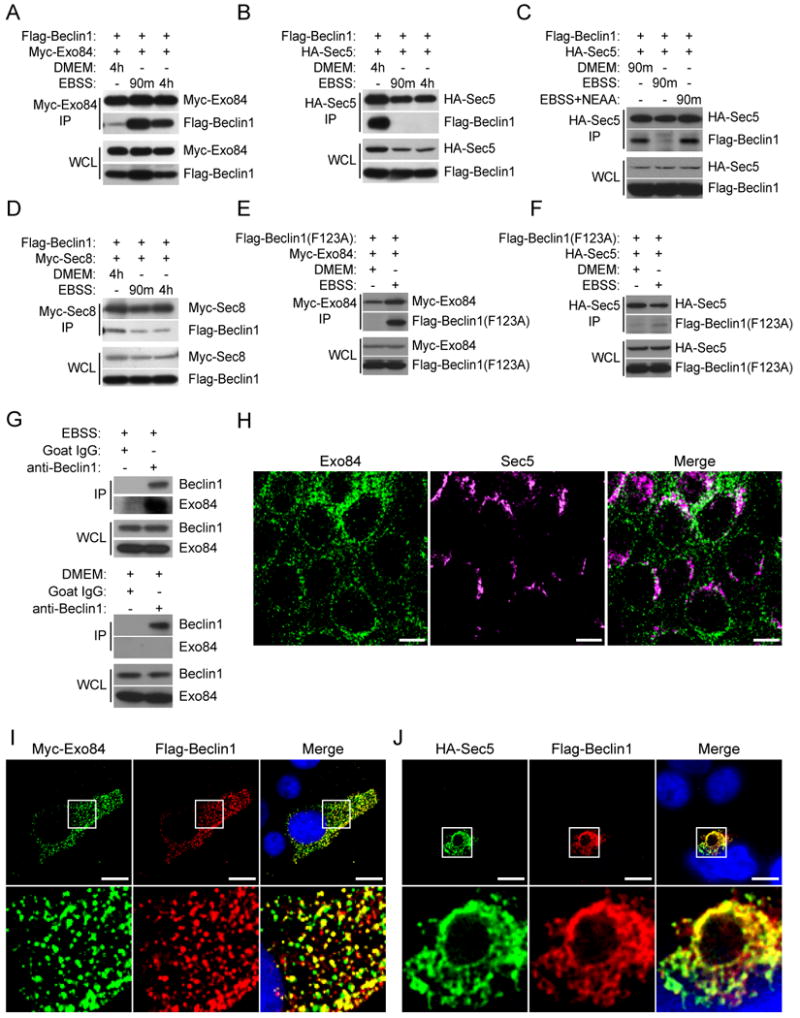
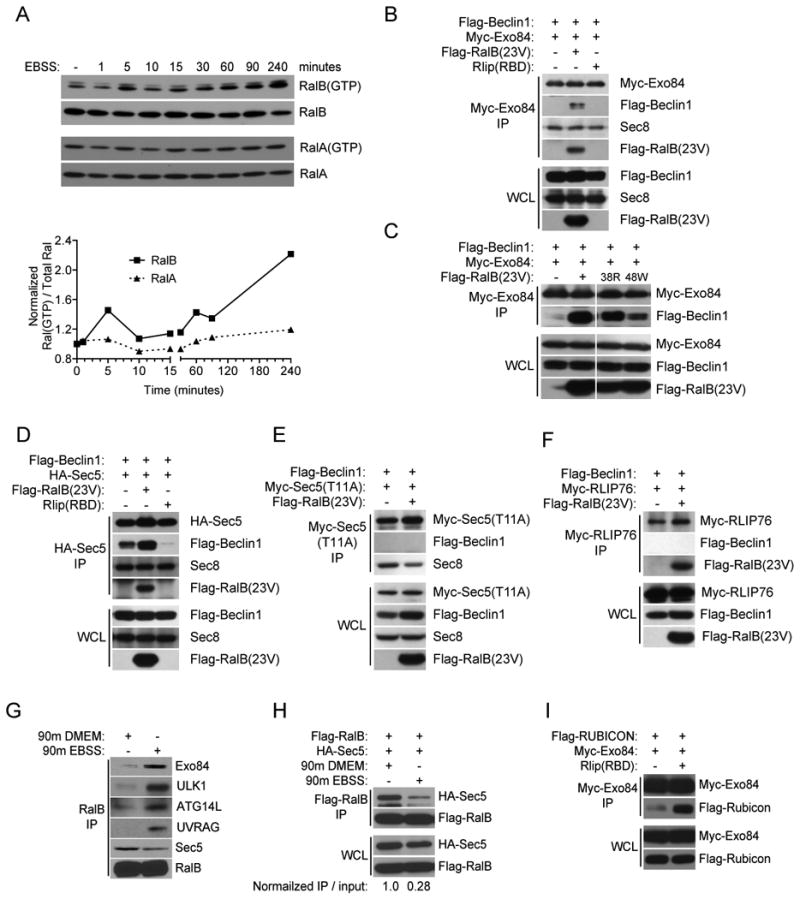
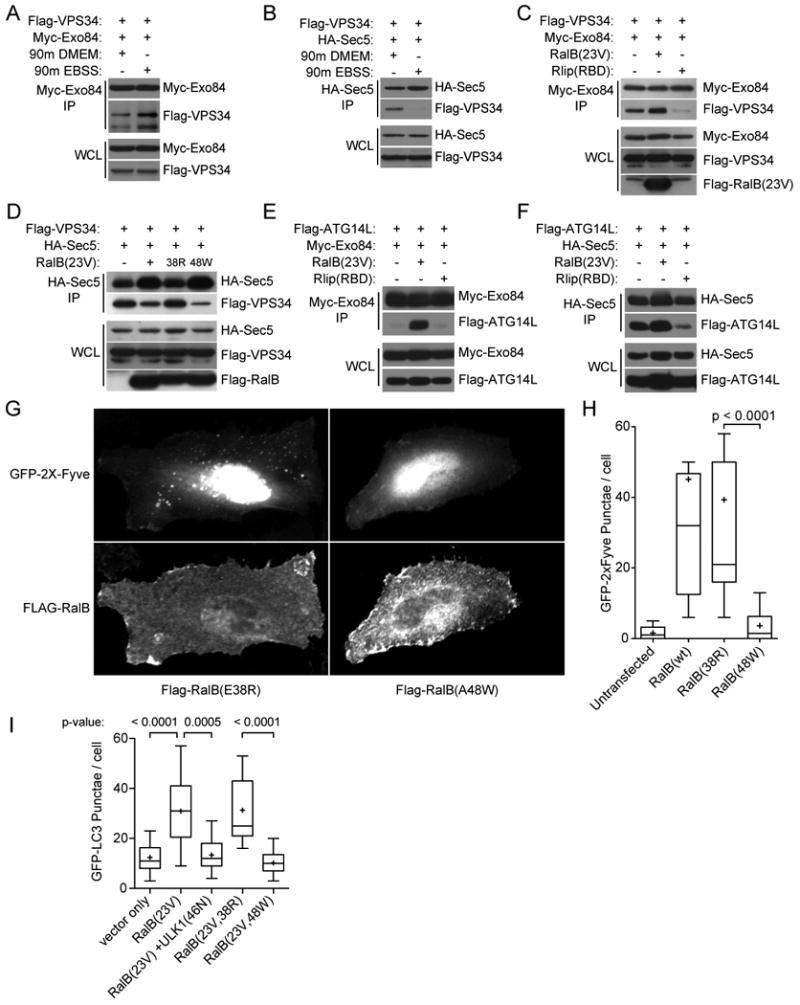
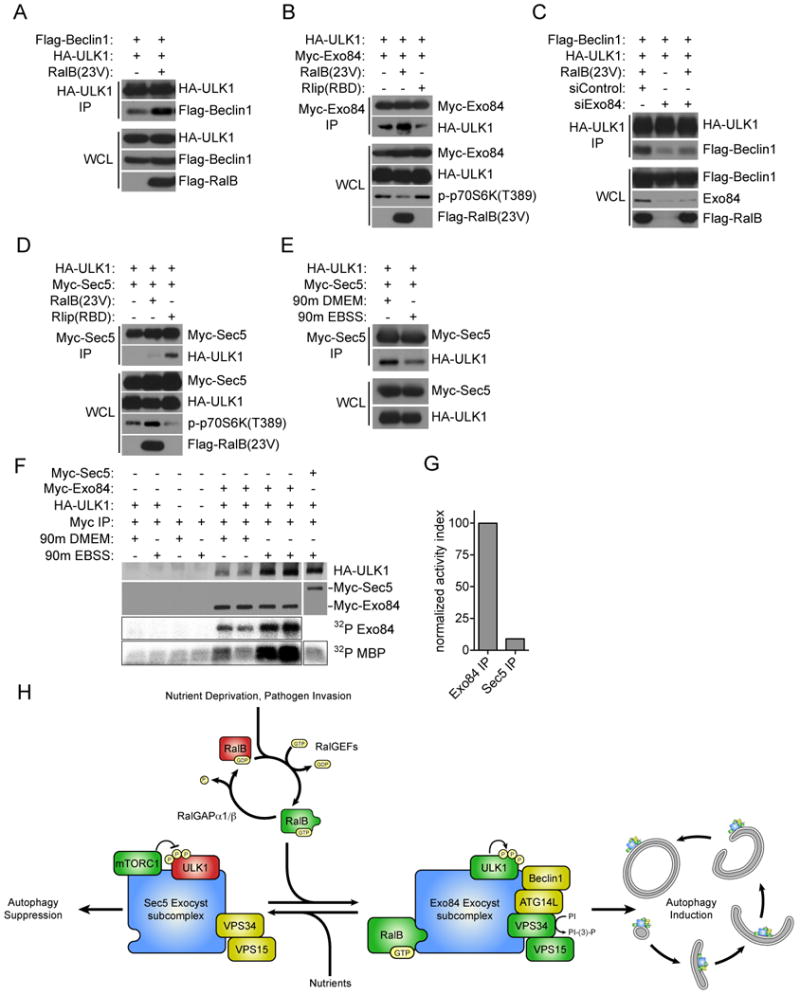
The study of macroautophagy in mammalian cells has described induction, vesicle nucleation, and membrane elongation complexes as key signaling intermediates driving autophagosome biogenesis. How these components are recruited to nascent autophagosomes is poorly understood, and although much is known about signaling mechanisms that restrain autophagy, the nature of positive inductive signals that can promote autophagy remain cryptic. We find that the Ras-like small G protein, RalB, is localized to nascent autophagosomes and is activated on nutrient deprivation. RalB and its effector Exo84 are required for nutrient starvation-induced autophagocytosis, and RalB activation is sufficient to promote autophagosome formation. Through direct binding to Exo84, RalB induces the assembly of catalytically active ULK1 and Beclin1-VPS34 complexes on the exocyst, which are required for isolation membrane formation and maturation. Thus, RalB signaling is a primary adaptive response to nutrient limitation that directly engages autophagocytosis through mobilization of the core vesicle nucleation machinery.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Rallying the exocyst as an autophagy scaffold.Cell. 2011 Jan 21;144(2):172-4. doi: 10.1016/j.cell.2011.01.005. Cell. 2011. PMID: 21241888 Free PMC article.
References
-
- Bhuvanakantham R, Li J, Tan TT, Ng ML. Human Sec3 protein is a novel transcriptional and translational repressor of flavivirus. Cell Microbiol. 2010;12:453–472. - PubMed
-
- Bodemann BO, White MA. Ral GTPases and cancer: linchpin support of the tumorigenic platform. Nat Rev Cancer. 2008;8:133–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
