

Mechanistic analysis of massive endocytosis in relation to functionally defined surface membrane domains
- PMID: 21242299
- PMCID: PMC3032373
- DOI: 10.1085/jgp.201010470
Mechanistic analysis of massive endocytosis in relation to functionally defined surface membrane domains
Abstract
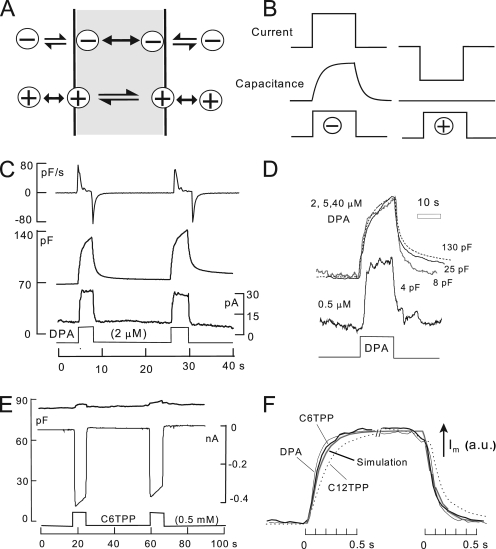
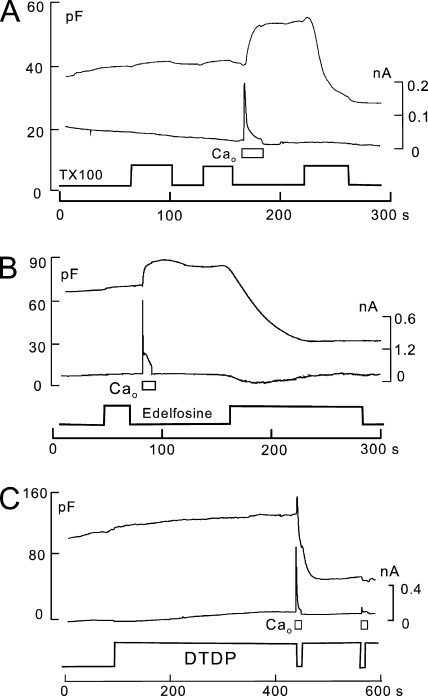
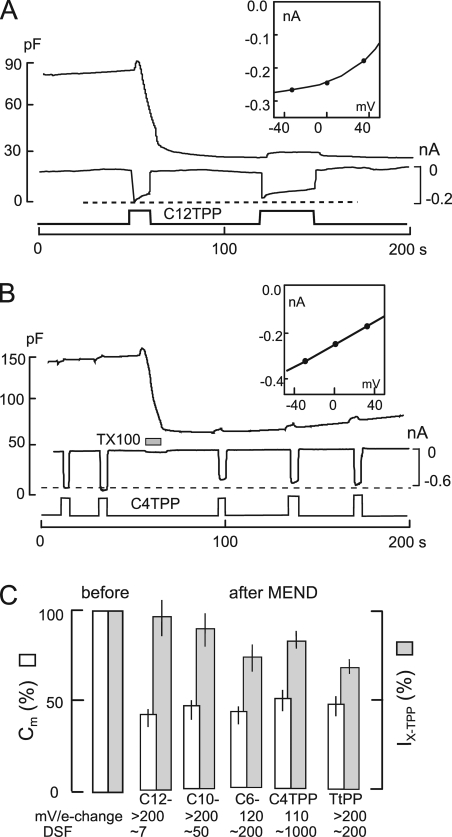
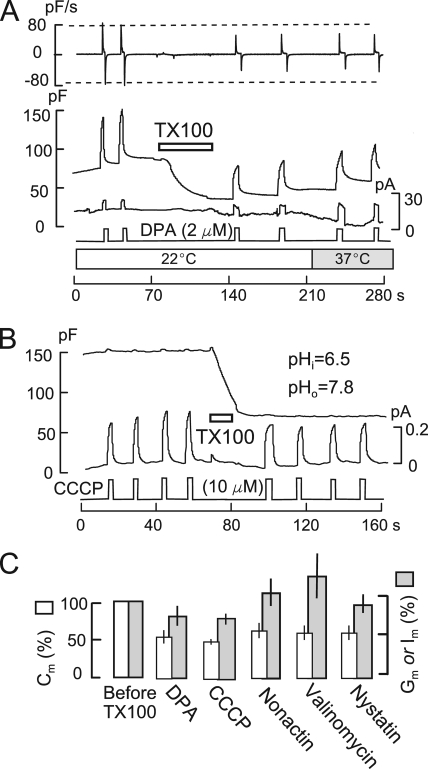
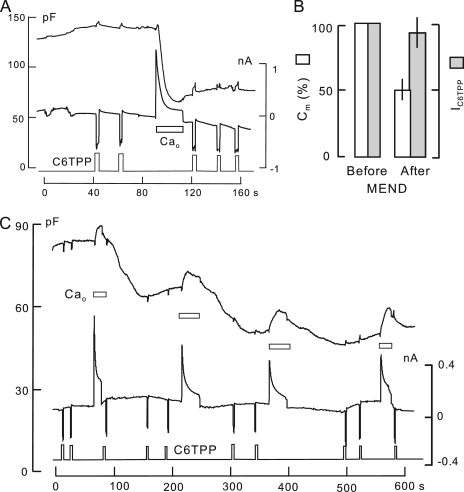
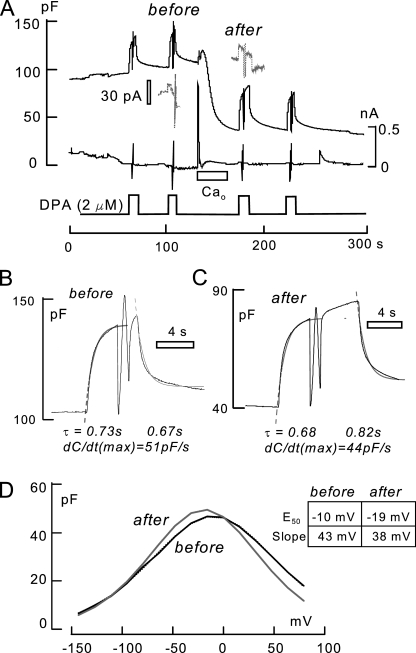
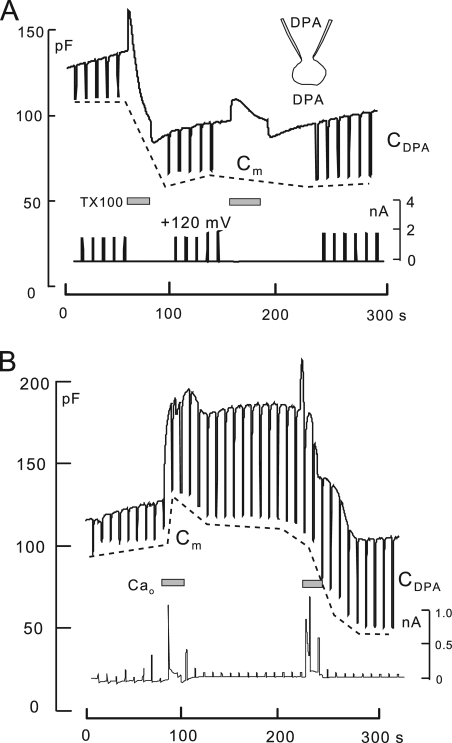
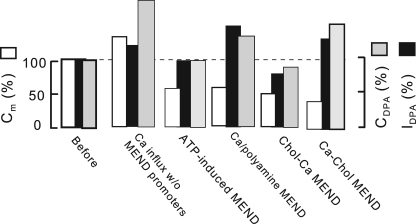
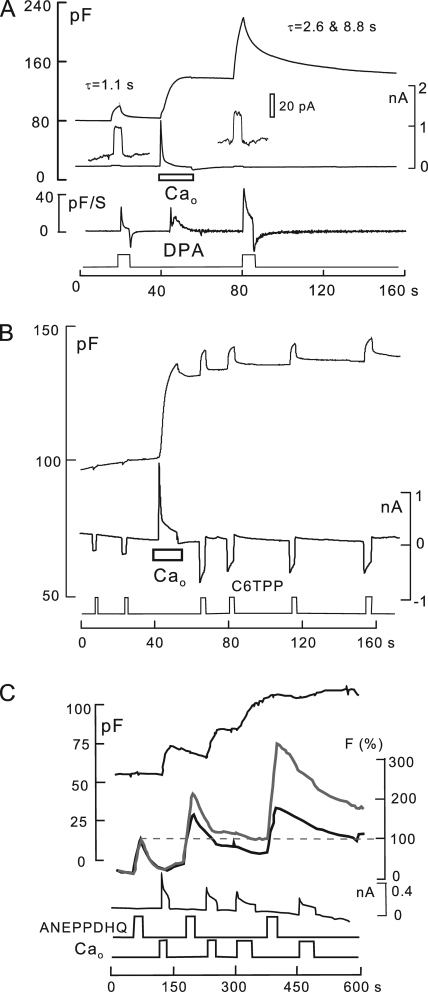
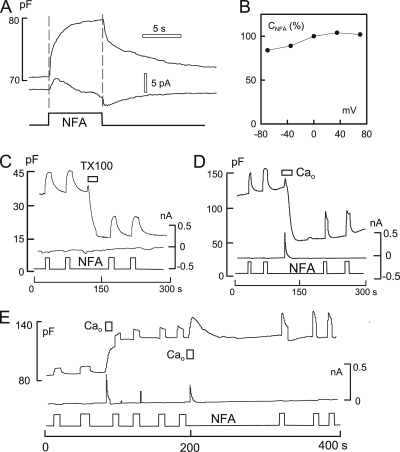
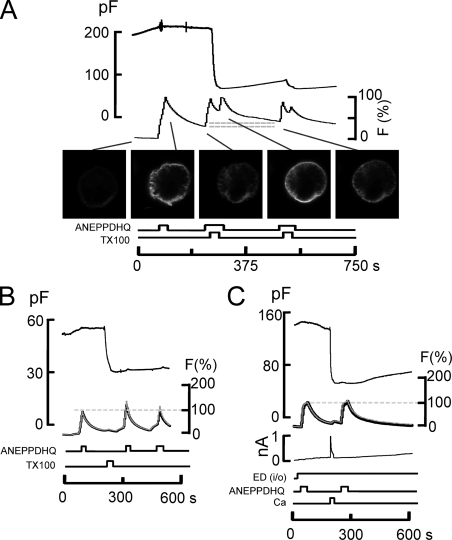
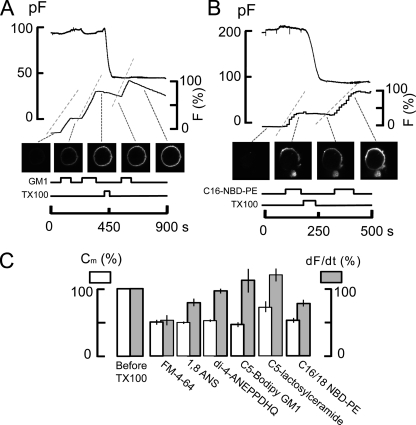
A large fraction of endocytosis in eukaryotic cells occurs without adaptors or dynamins. Here, we present evidence for the involvement of lipid domains in massive endocytosis (MEND) activated by both large Ca transients and amphipathic compounds in baby hamster kidney and HEK293 cells. First, we demonstrate functional coupling of the two MEND types. Ca transients can strongly facilitate detergent-activated MEND. Conversely, an amphipath with dual alkyl chains, ditridecylphthalate, is without effect in the absence of Ca transients but induces MEND to occur within seconds during Ca transients. Ca transients, like amphipaths, enhance the extraction of lipids from cells by β-cyclodextrins. Second, we demonstrate that electrical and/or optical signals generated by selected membrane probes are nearly insensitive to MEND, suggesting that those probes segregate into membrane domains that are not taken up by MEND. Triphenylphosphoniums are increasingly excluded from domains that internalize as the carbon chain length increases from 4 to 12. The small cationic membrane dye, FM 4-64, binds well to domains that internalize, whereas a closely related dye with a larger hydrophobic moiety, di-4-ANEPPDHQ (ANEPPDHQ) is excluded. Multiple carrier-type ionophores and a small amphipathic anion, niflumic acid, are also excluded. Probes with modest MEND sensitivity include the hydrophobic anion, dipicrylamine, carbonyl cyanide m-chlorophenylhydrazone, and NBD-phosphatidylethanolamine. Third, we demonstrate that large Ca transients can strongly enhance the extracellular binding of several membrane probes, monitored electrically or optically, consistent with a more disordered membrane with more amphipath-binding sites. Fluorescence shifts of ANEPPDHQ report increased disorder of the extracellular monolayer after large Ca transients, consistent with an increased propensity of the membrane to phase separate and vesiculate. Collectively, the results indicate that >50% of the outer monolayer is ordered and can be selectively internalized during MEND responses initiated by two very different cell perturbations.
Figures
Comment in
-
Cells on the MEND: exploring the role of lipidic forces in membrane trafficking.J Gen Physiol. 2011 Feb;137(2):133-6. doi: 10.1085/jgp.201010594. Epub 2011 Jan 17. J Gen Physiol. 2011. PMID: 21242298 Free PMC article. No abstract available.
References
-
- Brown D.A. 2006. Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology (Bethesda). 21:430–439 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
