

Elucidating the role of matrix stiffness in 3D cell migration and remodeling
- PMID: 21244824
- PMCID: PMC3021668
- DOI: 10.1016/j.bpj.2010.11.082
Elucidating the role of matrix stiffness in 3D cell migration and remodeling
Abstract
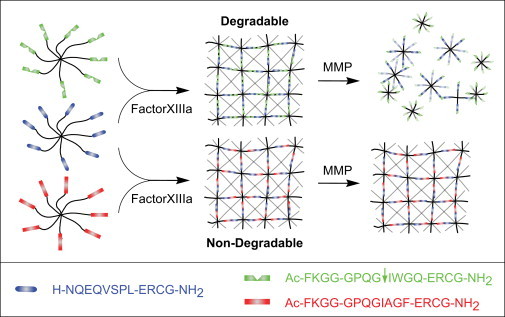
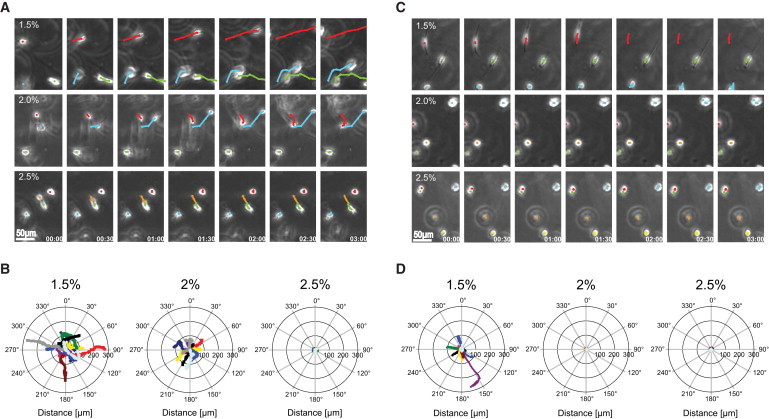
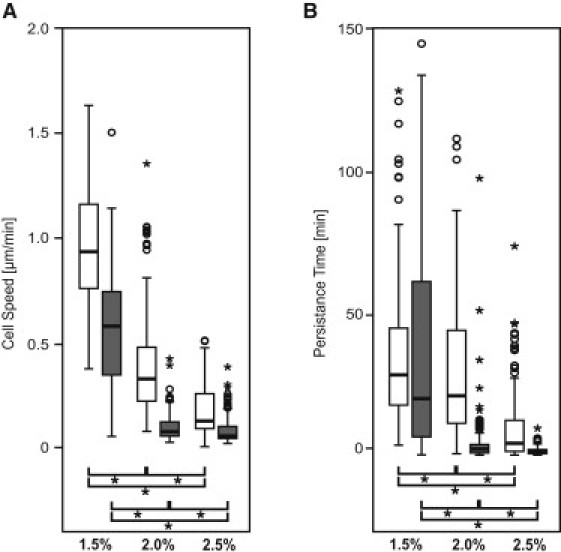
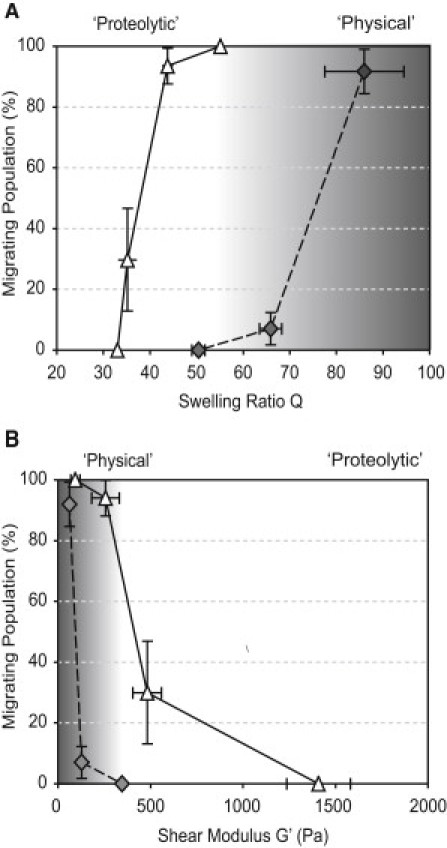
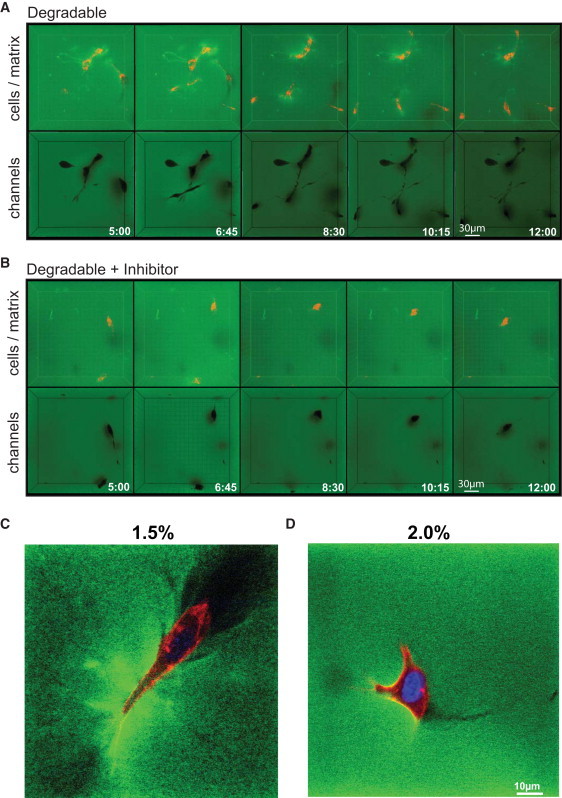
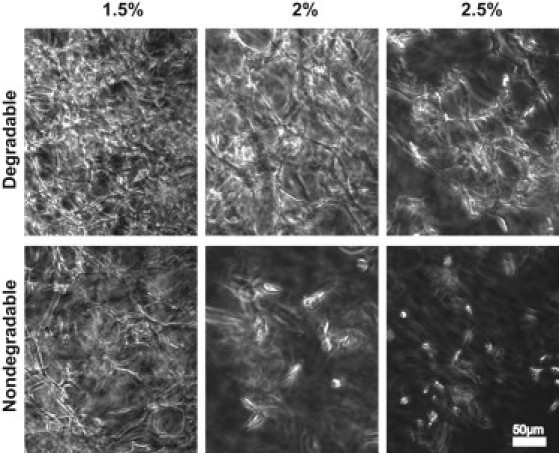
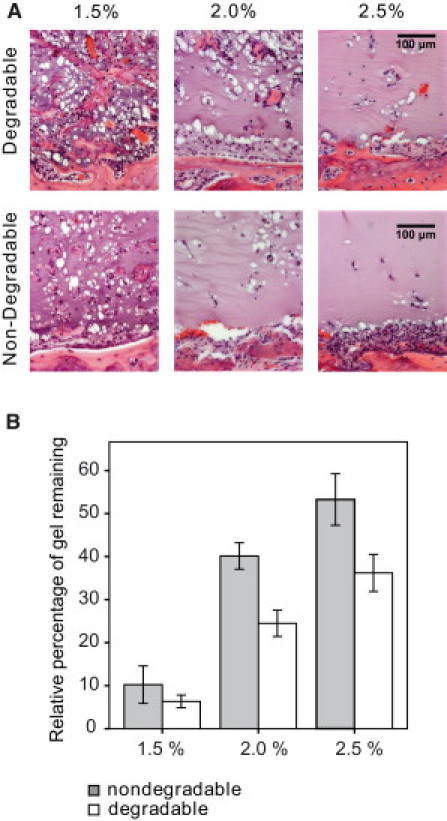
Reductionist in vitro model systems which mimic specific extracellular matrix functions in a highly controlled manner, termed artificial extracellular matrices (aECM), have increasingly been used to elucidate the role of cell-ECM interactions in regulating cell fate. To better understand the interplay of biophysical and biochemical effectors in controlling three-dimensional cell migration, a poly(ethylene glycol)-based aECM platform was used in this study to explore the influence of matrix cross-linking density, represented here by stiffness, on cell migration in vitro and in vivo. In vitro, the migration behavior of single preosteoblastic cells within hydrogels of varying stiffness and susceptibilities to degradation by matrix metalloproteases was assessed by time-lapse microscopy. Migration behavior was seen to be strongly dependent on matrix stiffness, with two regimes identified: a nonproteolytic migration mode dominating at relatively low matrix stiffness and proteolytic migration at higher stiffness. Subsequent in vivo experiments revealed a similar stiffness dependence of matrix remodeling, albeit less sensitive to the matrix metalloprotease sensitivity. Therefore, our aECM model system is well suited to unveil the role of biophysical and biochemical determinants of physiologically relevant cell migration phenomena.
Copyright © 2011 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Lutolf M.P., Hubbell J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005;23:47–55. - PubMed
-
- Mooney D.J., Vandenburgh H. Cell delivery mechanisms for tissue repair. Cell Stem Cell. 2008;2:205–213. - PubMed
-
- Zaman M.H., Matsudaira P., Lauffenburger D.A. Understanding effects of matrix protease and matrix organization on directional persistence and translational speed in three-dimensional cell migration. Ann. Biomed. Eng. 2007;35:91–100. - PubMed
-
- Ilina O., Friedl P. Mechanisms of collective cell migration at a glance. J. Cell Sci. 2009;122:3203–3208. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
