

Preferential accumulation of N-terminal mutant huntingtin in the nuclei of striatal neurons is regulated by phosphorylation
- PMID: 21245084
- PMCID: PMC3049362
- DOI: 10.1093/hmg/ddr023
Preferential accumulation of N-terminal mutant huntingtin in the nuclei of striatal neurons is regulated by phosphorylation
Abstract
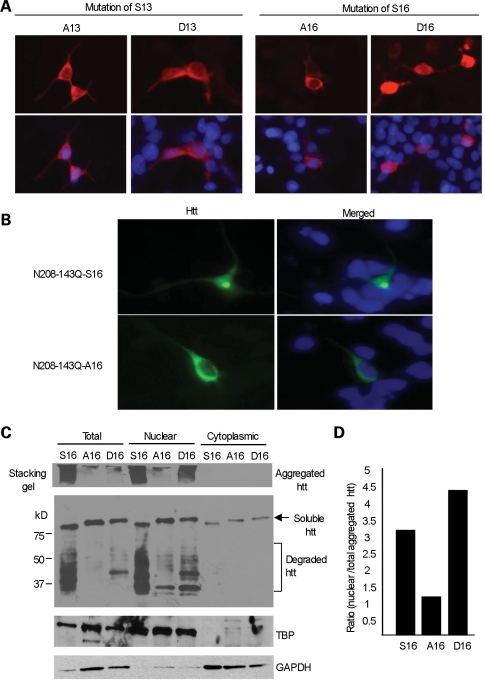
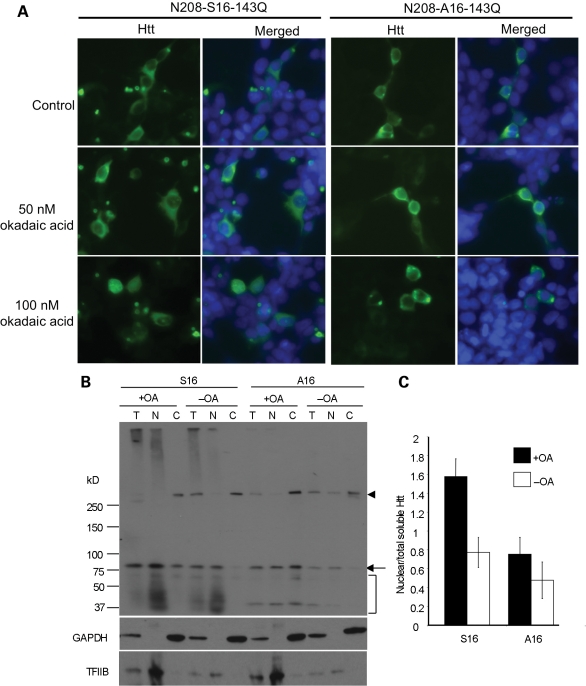
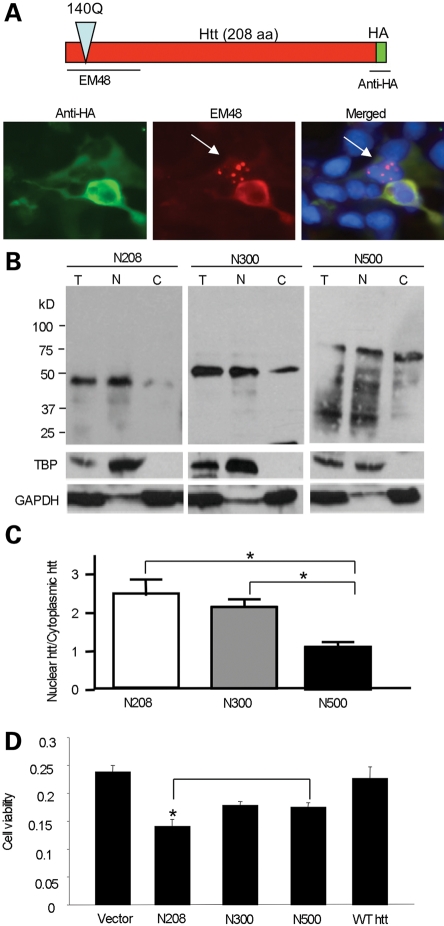
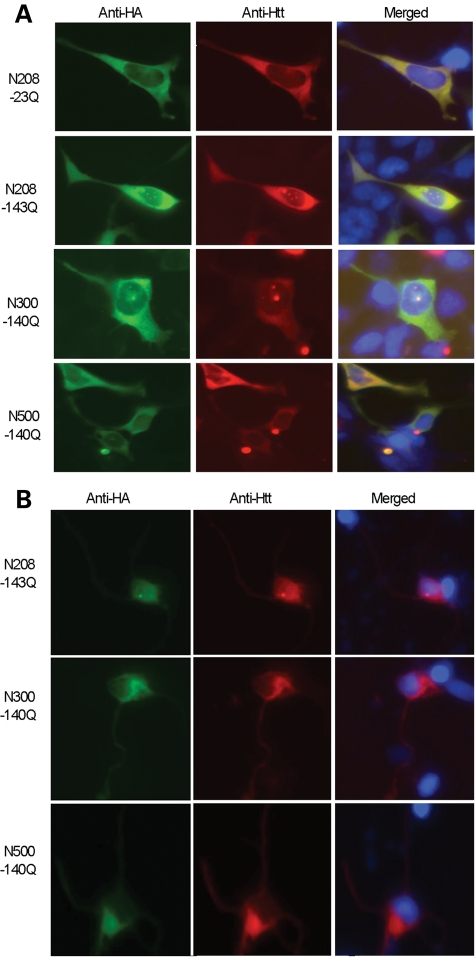
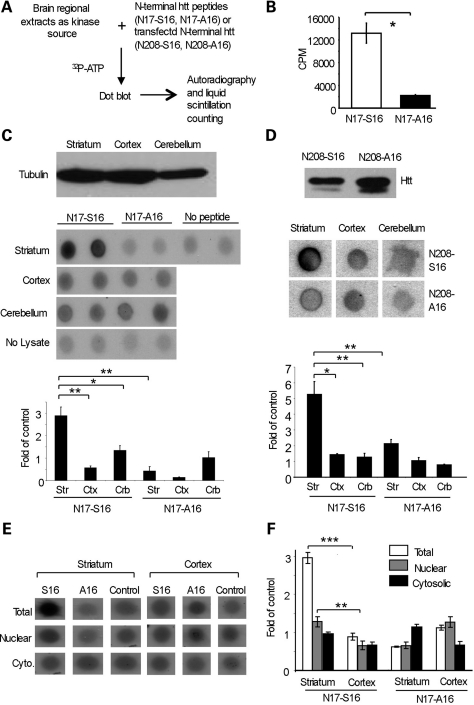
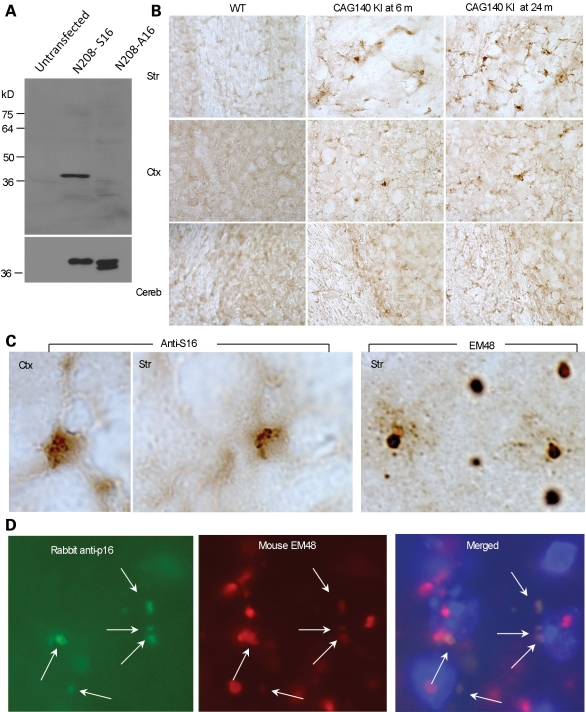
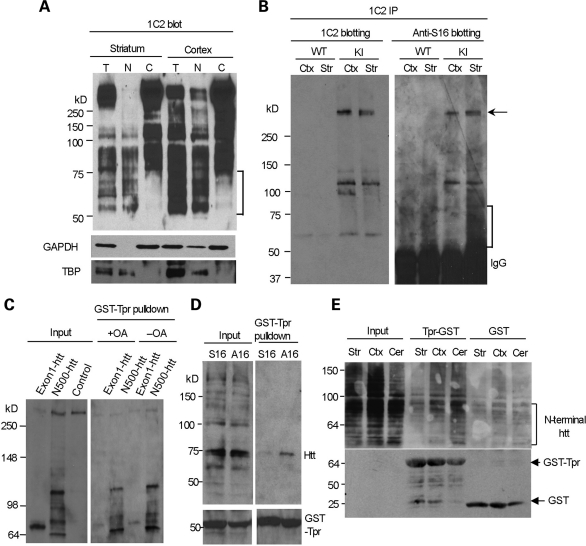
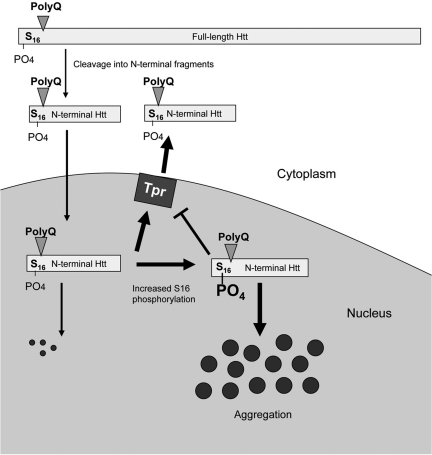
An expanded polyglutamine tract (>37 glutamines) in the N-terminal region of huntingtin (htt) causes htt to accumulate in the nucleus, leading to transcriptional dysregulation in Huntington disease (HD). In HD knock-in mice that express full-length mutant htt at the endogenous level, mutant htt preferentially accumulates in the nuclei of striatal neurons, which are affected most profoundly in HD. The mechanism underlying this preferential nuclear accumulation of mutant htt in striatal neurons remains unknown. Here, we report that serine 16 (S16) in htt is important for the generation of small N-terminal fragments that are able to accumulate in the nucleus and form aggregates. Phosphorylation of N-terminal S16 in htt promotes the nuclear accumulation of small N-terminal fragments and reduces the interaction of N-terminal htt with the nuclear pore complex protein Tpr. Mouse brain striatal tissues show increased S16 phosphorylation and a decreased association between mutant N-terminal htt and Tpr. These findings provide mechanistic insight into the nuclear accumulation of mutant htt and the selective neuropathology of HD, revealing potential therapeutic targets for treating this disease.
Figures
References
-
- DiFiglia M., Sapp E., Chase K.O., Davies S.W., Bates G.P., Vonsattel J.P., Aronin N. Aggregation of huntingtin in neuronal intranuclear inclusions and dystrophic neurites in brain. Science. 1997;277:1990–1993. doi:10.1126/science.277.5334.1990. - DOI - PubMed
-
- Orr H.T., Zoghbi H.Y. Trinucleotide repeat disorders. Annu. Rev. Neurosci. 2007;30:575–621. doi:10.1146/annurev.neuro.29.051605.113042. - DOI - PubMed
-
- Van Raamsdonk J.M., Murphy Z., Slow E.J., Leavitt B.R., Hayden M.R. Selective degeneration and nuclear localization of mutant huntingtin in the YAC128 mouse model of Huntington disease. Hum. Mol. Genet. 2005;14:3823–3835. doi:10.1093/hmg/ddi407. - DOI - PubMed
-
- Landles C., Bates G.P. Huntingtin and the molecular pathogenesis of Huntington's disease. Fourth in molecular medicine review series. EMBO Rep. 2004;5:958–963. doi:10.1038/sj.embor.7400250. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
