

Membrane surface charge dictates the structure and function of the epithelial Na+/H+ exchanger
- PMID: 21245831
- PMCID: PMC3041952
- DOI: 10.1038/emboj.2010.356
Membrane surface charge dictates the structure and function of the epithelial Na+/H+ exchanger
Abstract
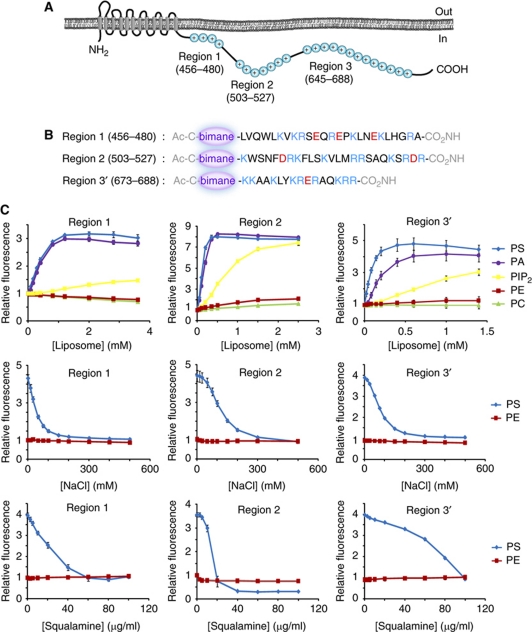
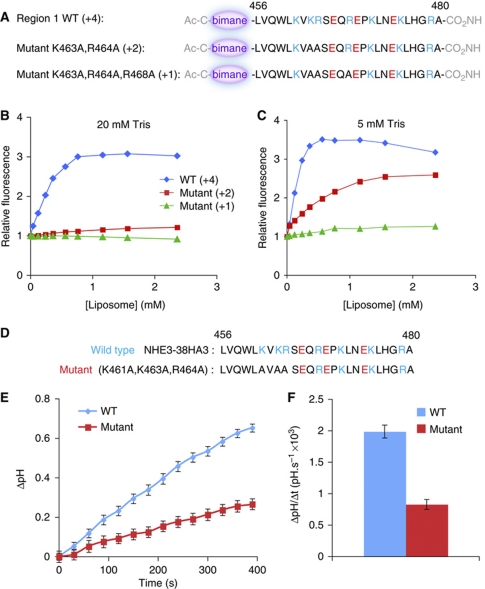
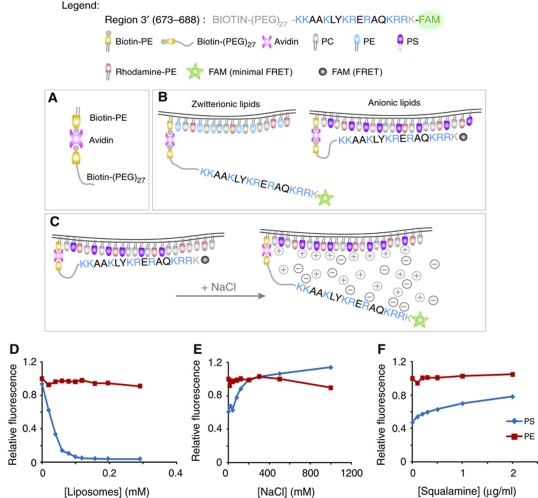
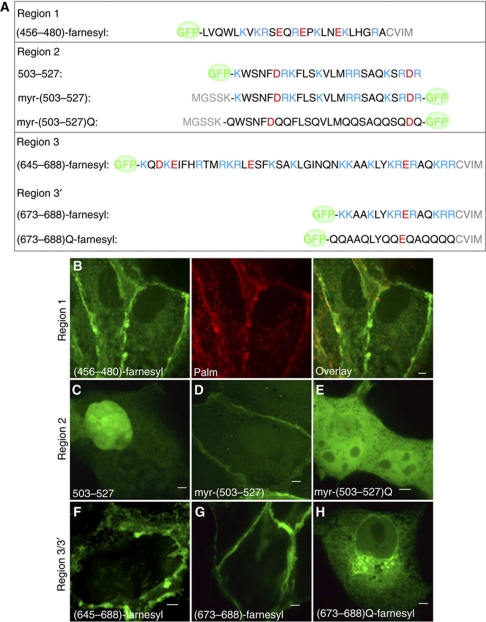
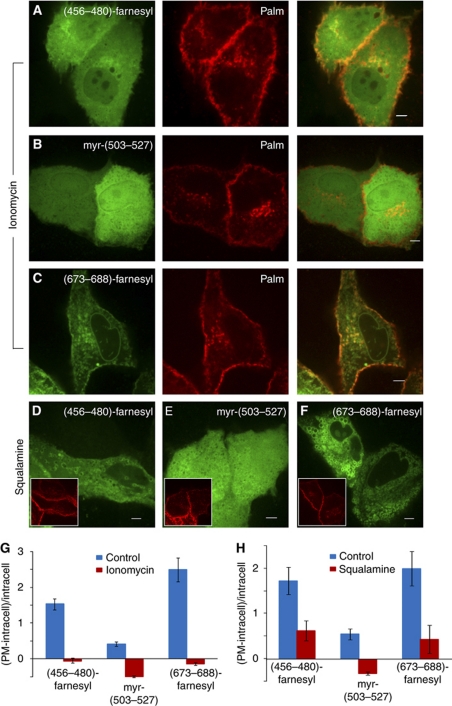
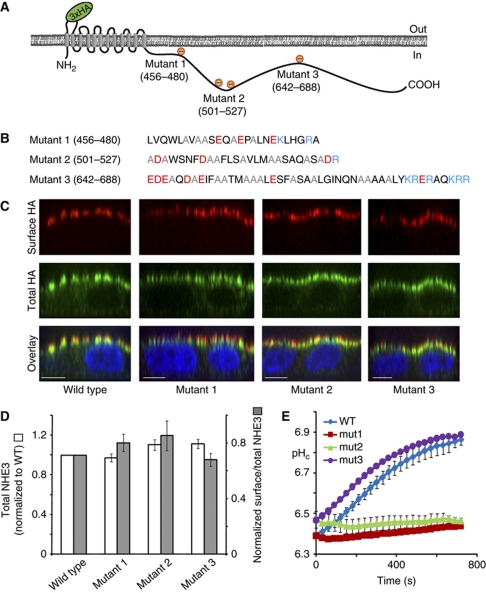
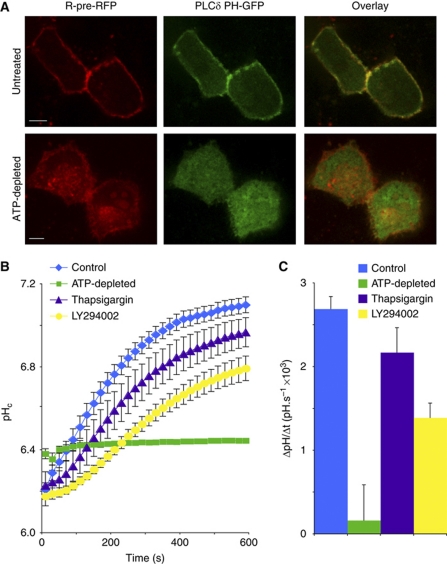
The Na(+)/H(+) exchanger NHE3 plays a central role in intravascular volume and acid-base homeostasis. Ion exchange activity is conferred by its transmembrane domain, while regulation of the rate of transport by a variety of stimuli is dependent on its cytosolic C-terminal region. Liposome- and cell-based assays employing synthetic or recombinant segments of the cytosolic tail demonstrated preferential association with anionic membranes, which was abrogated by perturbations that interfere with electrostatic interactions. Resonance energy transfer measurements indicated that segments of the C-terminal domain approach the bilayer. In intact cells, neutralization of basic residues in the cytosolic tail by mutagenesis or disruption of electrostatic interactions inhibited Na(+)/H(+) exchange activity. An electrostatic switch model is proposed to account for multiple aspects of the regulation of NHE3 activity.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Aharonovitz O, Demaurex N, Woodside M, Grinstein S (1999) ATP dependence is not an intrinsic property of Na+/H+ exchanger NHE1: requirement for an ancillary factor. Am J Physiol 276: C1303–C1311 - PubMed
-
- Akhter S, Cavet ME, Tse CM, Donowitz M (2000) C-terminal domains of Na+/H+ exchanger isoform 3 are involved in the basal and serum-stimulated membrane trafficking of the exchanger. Biochemistry 39: 1990–2000 - PubMed
-
- Akhter S, Nath SK, Tse CM, Williams J, Zasloff M, Donowitz M (1999) Squalamine, a novel cationic steroid, specifically inhibits the brush-border Na+/H+ exchanger isoform NHE3. Am J Physiol 276: C136–C144 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
