

Review
doi: 10.1002/dvdy.22540.
Epub 2011 Jan 18.
Vertebrate intestinal endoderm development
- PMID: 21246663
- PMCID: PMC3079549
- DOI: 10.1002/dvdy.22540
Item in Clipboard
Review
Vertebrate intestinal endoderm development
Dev Dyn.
2011 Mar.
Abstract
The endoderm gives rise to the lining of the esophagus, stomach and intestines, as well as associated organs. To generate a functional intestine, a series of highly orchestrated developmental processes must occur. In this review, we attempt to cover major events during intestinal development from gastrulation to birth, including endoderm formation, gut tube growth and patterning, intestinal morphogenesis, epithelial reorganization, villus emergence, as well as proliferation and cytodifferentiation. Our discussion includes morphological and anatomical changes during intestinal development as well as molecular mechanisms regulating these processes.
Copyright © 2011 Wiley-Liss, Inc.
Figures

A. Schematic of the morphogenetic movements giving rise to the gut tube from post-gastrulation (E7.25) through gut tube formation (E9.5) in the mouse embryo. At E7.25, after gastrulation, the embryo is organized in a cup shape with the endoderm on the outer-most surface (yellow) and the mesoderm and ectoderm on the inside of the cup (black). At E7.75, pits that will give rise to the AIP and CIP are evident (black arrows). Note that the AIP forms several hours prior to the CIP, but for ease of illustration they are shown together. By E8.0 the AIP and CIP are clearly visible. Between E8.0 and E9.0, the lip of both the portals moves toward the center of the embryo, and the lateral endoderm continues to fold ventrally. Between E9.0 and E9.5 the endoderm has finished folding and exists as a tube surrounded by mesenchyme. B. Ventral view of the endoderm folding. At E7.5, shortly after gastrulation has ended, the endoderm is schematically represented as a relatively naive flat sheet. By E7.75, the endoderm begins to fold along the longitudinal axis, much like a sheet of paper being rolled into a tube. At the same time, the intestinal portals begin to form (black arrows point to invaginations where the portals will form). Between E8.0 and E8.5, the ends of the endodermal “tube” are formed and represent the anterior and caudal intestinal portals. Simultaneously, the endoderm continues to fold ventrally along the longitudinal axis. Since the endoderm is covered by mesoderm, as the endoderm forms a tube, mesoderm is visible. By E9.0–E9.5 the endodermal tube is closed such that the endoderm is entirely surrounded by mesoderm. The dashed line denotes a cross section through the closed tube, where the endoderm is surrounded by the mesoderm. C. FoxA2creER;R26R embryos treated with Tamoxifen via maternal oral gavage (0.12mg/g) at E6.5. Tamoxifen activation of FoxA2creER causes extensive recombination of the R26R allele, leading to β-galactosidase expression in the endoderm and notochord. Embryos were collected and stained for β-galactosidase activity with X-Gal (shown in blue) at E8.5 and E9.5. Anterior is denoted by the red arrow and posterior is denoted by the blue arrow. The middle panel demonstrates the CIP at E8.5, with the arrow pointing to the lip of the CIP.

A–B. At left, a schematic drawing of the embryo shows the level of the histological sections shown at right. The endoderm in colored yellow in the schematic and pseudocolored yellow in the histological sections. A. Transverse sections through the caudal region of an E7.75 embryo demonstrating the open endoderm mid-embryo (section 1) which is starting to form a CIP (section 2 and 3). B. Transverse sections through the caudal region of an E8.5 embryo showing the open gut tube toward the middle of the embryo, which closes forming the CIP at the posterior of the embryo. C. Changes in the gut tube epithelium between E9.0 and E11.5. The gut tube endoderm condenses between E9.0 and E9.5 to give rise to a pseudostratified epithelium. Between E9.5 and E10.5 the gut tube epithelium circumference increases giving rise to a bigger lumen and an increased epithelial surface area. All images are the same magnification (40X). Endoderm is pseudocolored yellow in all histological sections.
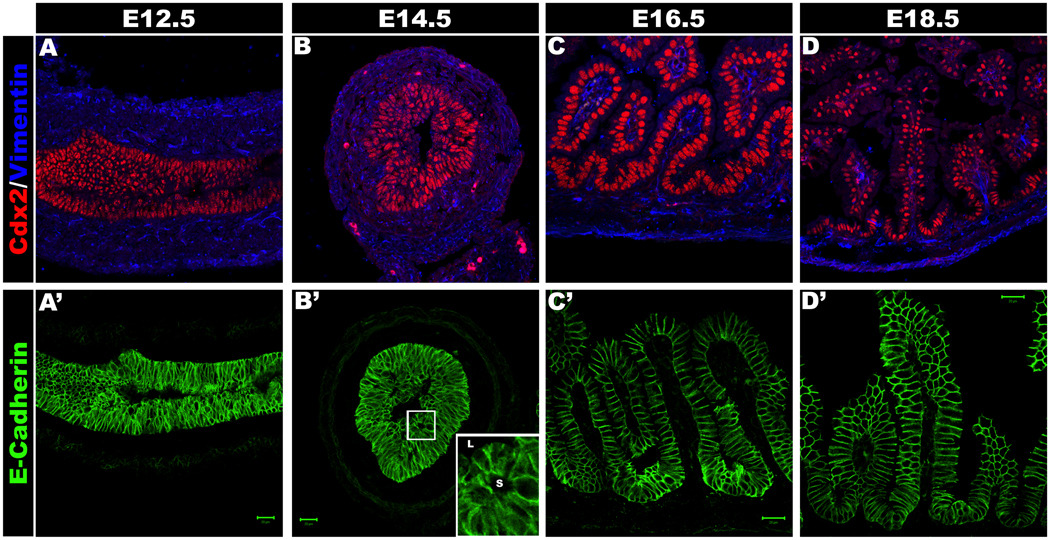
Intestinal development at E12.5 (A, A’), E14.5 (B, B’), E16.5 (C, C’) and E18.5 (D, D’). A–D shows the Cdx2 positive epithelium (Red) and Vimentin positive mesenchyme. A’–D’ shows the E-cadherin positive epithelium. Secondary lumina can be seen at E14.5 (Boxed region is magnified in the inset; L indicates primary lumen, s indicates secondary lumen) (B’) as epithelial reorganization begins. By E16.5 there are clear intervillus regions and villi (C, C’).

A. The grey box contains a schematic of an E8.0 mouse embryo with the endoderm in yellow. Signals involved in anterior-posterior patterning are sent from the mesoderm (Meso, white cells) to the endoderm (Endo, yellow cells). Signaling pathways important for A-P patterning are summarized, and include fine regulation of Bmp, Wnt, Fgf and RA signaling along the axis. B–F. The grey boxes contain a schematic of a transverse section through the proximal intestine (duodenum) at different developmental time points. The endoderm-derived intestinal epithelium is shown in yellow. A close up of the intestinal epithelium and underlying mesoderm-derived mesenchyme is shown under the grey box. Key molecular events regulating intestine development are shown based on the developmental time during which evidence has been presented for each event. B. Pseudostratified intestinal endoderm at E10.5. C. At E14.5, intestinal morphogenesis is underway. The intestinal epithelium becomes stratified and the formation of Ezrin+ secondary lumina is apparent (Ezrin expression is shown in red). D. By E15, secondary lumina have fused with the luminal surface and villus emergence is evident. In addition, condensed mesenchyme expressing higher levels of BMPs and PDGFR-α underlying nascent villi are present. Major signaling pathways (described in detail in the text) involved in epithelial-mesenchymal crosstalk responsible for modulating villus emergence are depicted. E. At E16.5 villi and intervillus regions are evident. Sox9 expression is restricted to the proliferative intervillus region (green cells). Recent evidence has shown that β-catenin activity is present in the villus epithelium, but is excluded from the proliferative intervillus region. F. By late E18.5, β-catenin activity has transitioned and is present in the proliferative Sox9+ intervillus villus, and is absent from the villi. A schematic of the pathways involved in mesenchymal-epithelial crosstalk at E18.5 that regulate both mesenchymal and epithelial proliferation is shown.
References
-
- Allan D, Houle M, Bouchard N, Meyer BI, Gruss P, Lohnes D. RARgamma and Cdx1 interactions in vertebral patterning. Dev Biol. 2001;240:46–60. - PubMed
-
- Ameri J, Stahlberg A, Pedersen J, Johansson JK, Johannesson MM, Artner I, Semb H. FGF2 specifies hESC-derived definitive endoderm into foregut/midgut cell lineages in a concentration-dependent manner. Stem Cells. 2010;28:45–56. - PubMed
-
- Aoki TO, David NB, Minchiotti G, Saint-Etienne L, Dickmeis T, Persico GM, Strahle U, Mourrain P, Rosa FM. Molecular integration of casanova in the Nodal signalling pathway controlling endoderm formation. Development. 2002;129:275–286. - PubMed
-
- Aubin J, Lemieux M, Tremblay M, Berard J, Jeannotte L. Early postnatal lethality in Hoxa-5 mutant mice is attributable to respiratory tract defects. Dev Biol. 1997;192:432–445. - PubMed
-
- Auclair BA, Benoit YD, Rivard N, Mishina Y, Perreault N. Bone morphogenetic protein signaling is essential for terminal differentiation of the intestinal secretory cell lineage. Gastroenterology. 2007;133:887–896. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
