

Proprotein convertases process Pmel17 during secretion
- PMID: 21247888
- PMCID: PMC3059051
- DOI: 10.1074/jbc.M110.168088
Proprotein convertases process Pmel17 during secretion
Abstract
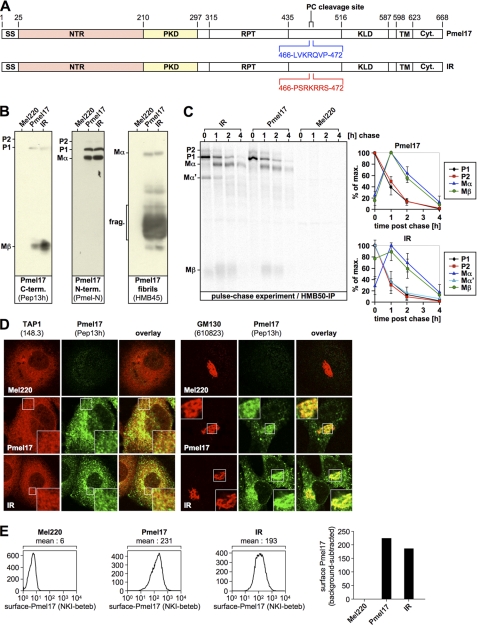
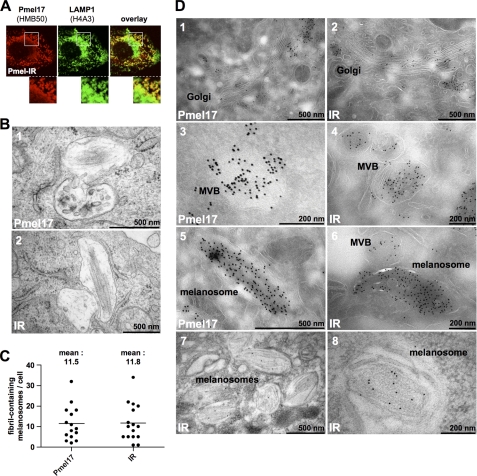
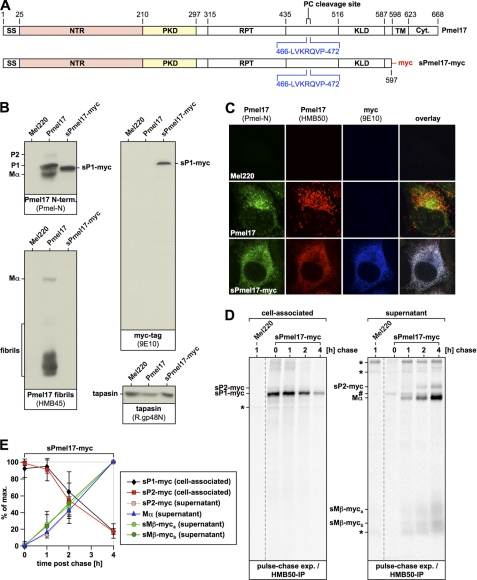
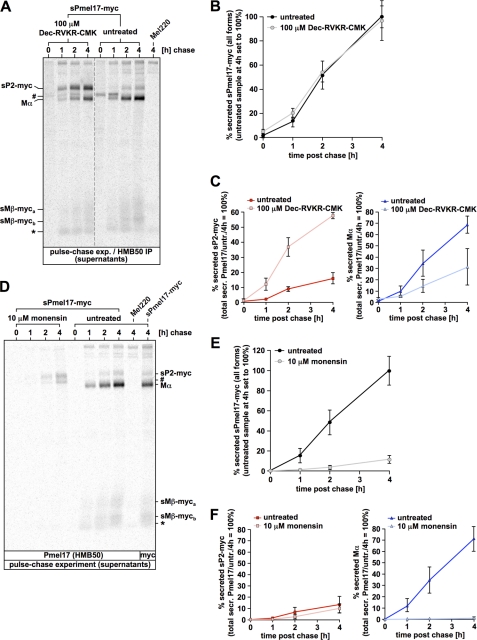
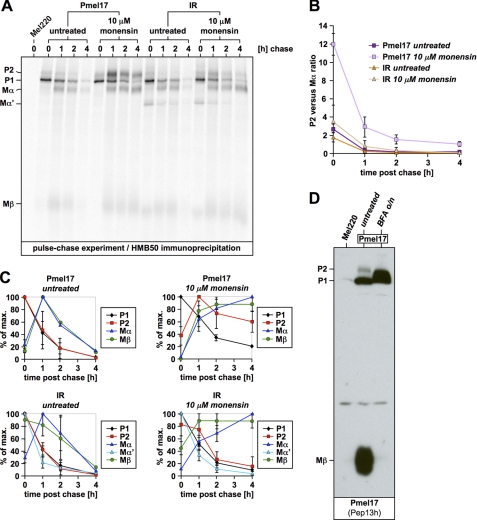
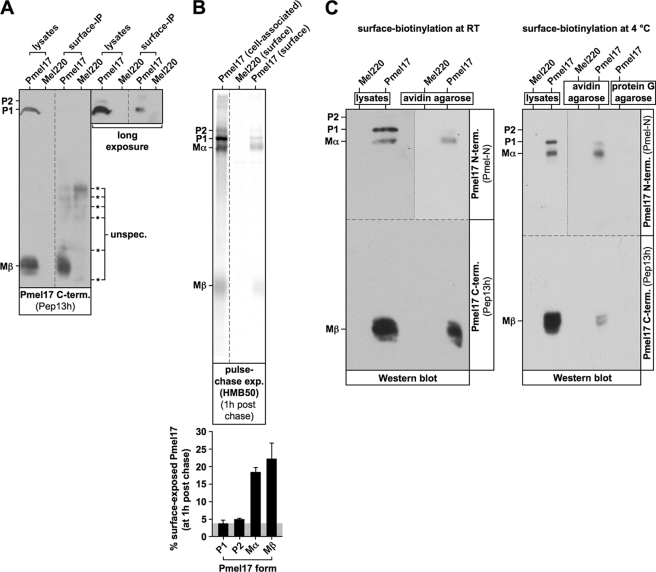
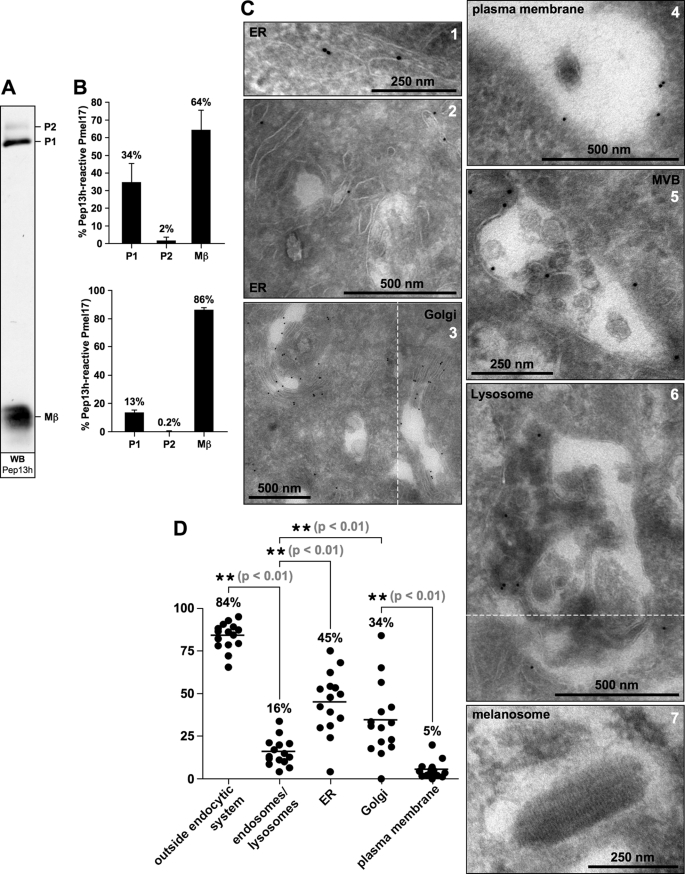
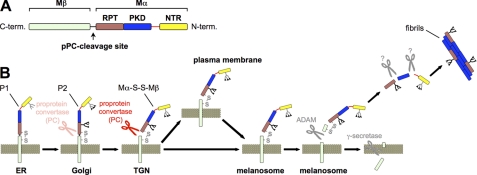
Pmel17 is a melanocyte/melanoma-specific protein that traffics to melanosomes where it forms a fibrillar matrix on which melanin gets deposited. Before being cleaved into smaller fibrillogenic fragments the protein undergoes processing by proprotein convertases, a class of serine proteases that typically recognize the canonical motif RX(R/K)R↓. The current model of Pmel17 maturation states that this processing step occurs in melanosomes, but in light of recent reports this issue has become controversial. We therefore addressed this question by thoroughly assessing the processing kinetics of either wild-type Pmel17 or a secreted soluble Pmel17 derivative. Our results demonstrate clearly that processing of Pmel17 occurs during secretion and that it does not require entry of the protein into the endocytic system. Strikingly, processing proceeds even in the presence of the secretion inhibitor monensin, suggesting that Pmel17 is an exceptionally good substrate. In line with this, we find that newly synthesized surface Pmel17 is already quantitatively cleaved. Moreover, we demonstrate that Pmel17 function is independent of the sequence identity of its unconventional proprotein convertase-cleavage motif that lacks arginine in P4 position. The data alter the current view of Pmel17 maturation and suggest that the multistep processing of Pmel17 begins with an early cleavage during secretion that primes the protein for later functional processing.
Figures
References
-
- Lee Z. H., Hou L., Moellmann G., Kuklinska E., Antol K., Fraser M., Halaban R., Kwon B. S. (1996) J. Invest. Dermatol. 106, 605–610 - PubMed
-
- Quevedo W. C., Fleischmann R. D., Dyckman J. (1981) in Phenotypic Expression in Pigment Cells (Seiji M. ed) pp. 177–184, Tokyo University Press, Tokyo
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
