

Identification of distal cis-regulatory elements at mouse mitoferrin loci using zebrafish transgenesis
- PMID: 21248200
- PMCID: PMC3135305
- DOI: 10.1128/MCB.01010-10
Identification of distal cis-regulatory elements at mouse mitoferrin loci using zebrafish transgenesis
Abstract
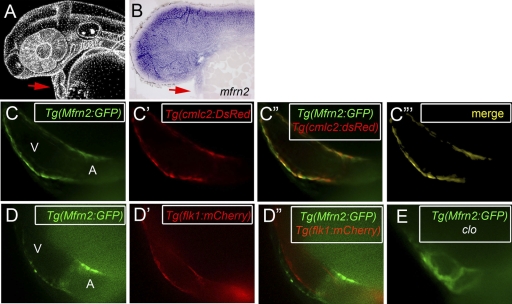
Mitoferrin 1 (Mfrn1; Slc25a37) and mitoferrin 2 (Mfrn2; Slc25a28) function as essential mitochondrial iron importers for heme and Fe/S cluster biogenesis. A genetic deficiency of Mfrn1 results in a profound hypochromic anemia in vertebrate species. To map the cis-regulatory modules (CRMs) that control expression of the Mfrn genes, we utilized genome-wide chromatin immunoprecipitation (ChIP) datasets for the major erythroid transcription factor GATA-1. We identified the CRMs that faithfully drive the expression of Mfrn1 during blood and heart development and Mfrn2 ubiquitously. Through in vivo analyses of the Mfrn-CRMs in zebrafish and mouse, we demonstrate their functional and evolutionary conservation. Using knockdowns with morpholinos and cell sorting analysis in transgenic zebrafish embryos, we show that GATA-1 directly regulates the expression of Mfrn1. Mutagenesis of individual GATA-1 binding cis elements (GBE) demonstrated that at least two of the three GBE within this CRM are functionally required for GATA-mediated transcription of Mfrn1. Furthermore, ChIP assays demonstrate switching from GATA-2 to GATA-1 at these elements during erythroid maturation. Our results provide new insights into the genetic regulation of mitochondrial function and iron homeostasis and, more generally, illustrate the utility of genome-wide ChIP analysis combined with zebrafish transgenesis for identifying long-range transcriptional enhancers that regulate tissue development.
Figures









References
-
- Bajic V. B., Tan S. L., Suzuki Y., Sugano S. 2004. Promoter prediction analysis on the whole human genome. Nat. Biotechnol. 22:1467–1473 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous