

Action potentials drive body wall muscle contractions in Caenorhabditis elegans
- PMID: 21248227
- PMCID: PMC3038695
- DOI: 10.1073/pnas.1012346108
Action potentials drive body wall muscle contractions in Caenorhabditis elegans
Abstract
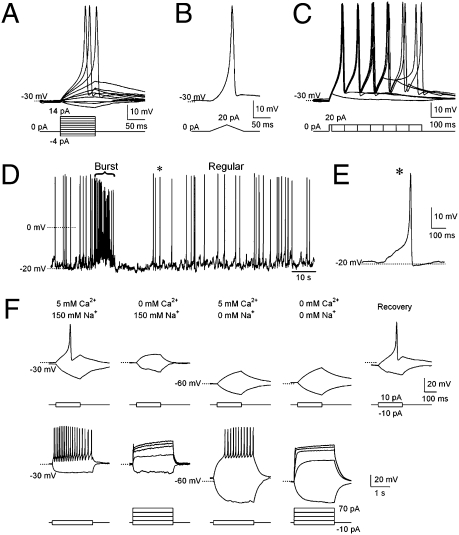
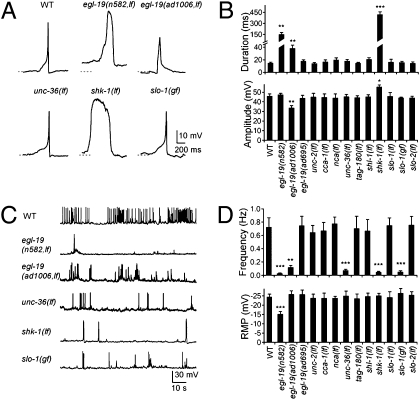
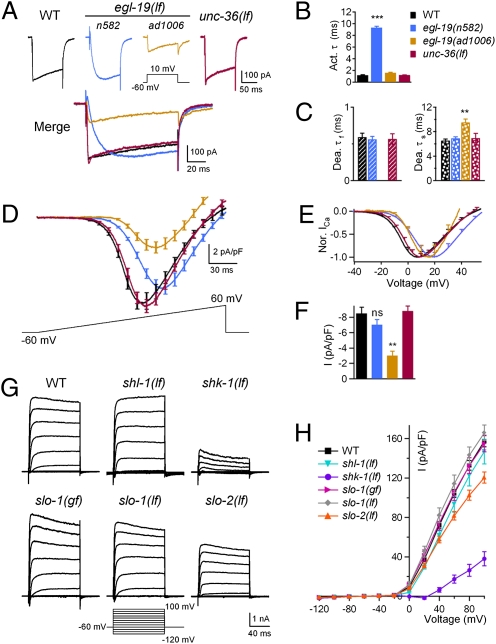
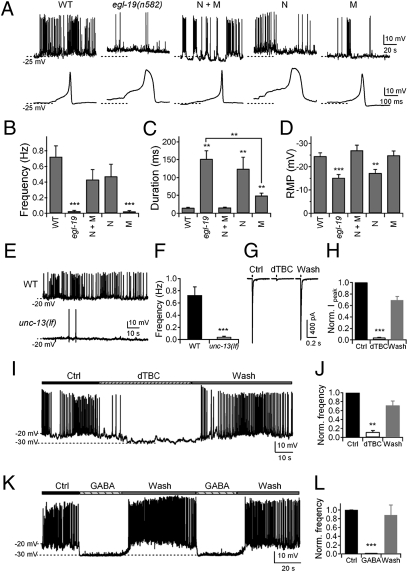
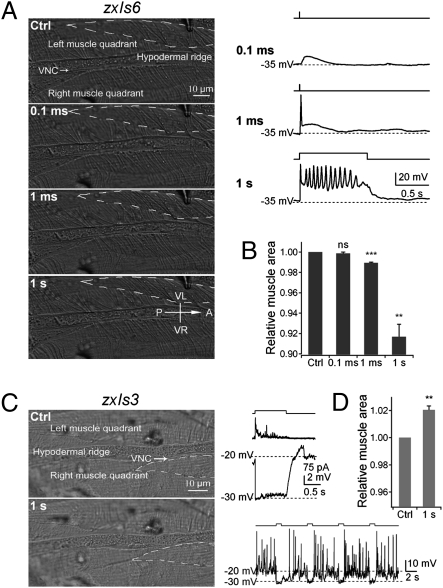
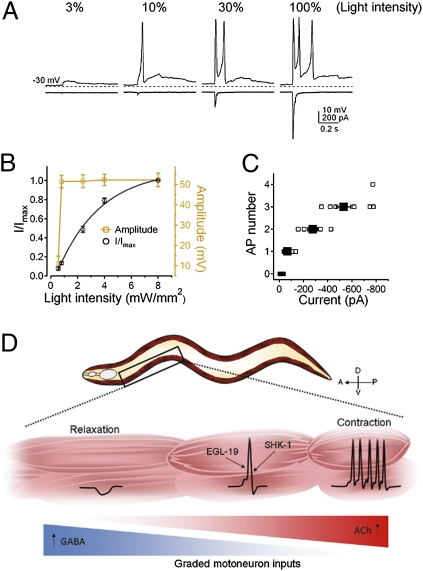
The sinusoidal locomotion exhibited by Caenorhabditis elegans predicts a tight regulation of contractions and relaxations of its body wall muscles. Vertebrate skeletal muscle contractions are driven by voltage-gated sodium channel-dependent action potentials. How coordinated motor outputs are regulated in C. elegans, which does not have voltage-gated sodium channels, remains unknown. Here, we show that C. elegans body wall muscles fire all-or-none, calcium-dependent action potentials that are driven by the L-type voltage-gated calcium and Kv1 voltage-dependent potassium channels. We further demonstrate that the excitatory and inhibitory motoneuron activities regulate the frequency of action potentials to coordinate muscle contraction and relaxation, respectively. This study provides direct evidence for the dual-modulatory model of the C. elegans motor circuit; moreover, it reveals a mode of motor control in which muscle cells integrate graded inputs of the nervous system and respond with all-or-none electrical signals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sulston JE, Schierenberg E, White JG, Thomson JN. The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev Biol. 1983;100:64–119. - PubMed
-
- White JG, Southgate E, Thomson JN, Brenner S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos Trans R Soc Lond B Biol Sci. 1986;314:1–340. - PubMed
-
- Bargmann CI. Neurobiology of the Caenorhabditis elegans genome. Science. 1998;282:2028–2033. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
