

Large duplications at reciprocal translocation breakpoints that might be the counterpart of large deletions and could arise from stalled replication bubbles
- PMID: 21252201
- PMCID: PMC3065700
- DOI: 10.1101/gr.114116.110
Large duplications at reciprocal translocation breakpoints that might be the counterpart of large deletions and could arise from stalled replication bubbles
Abstract
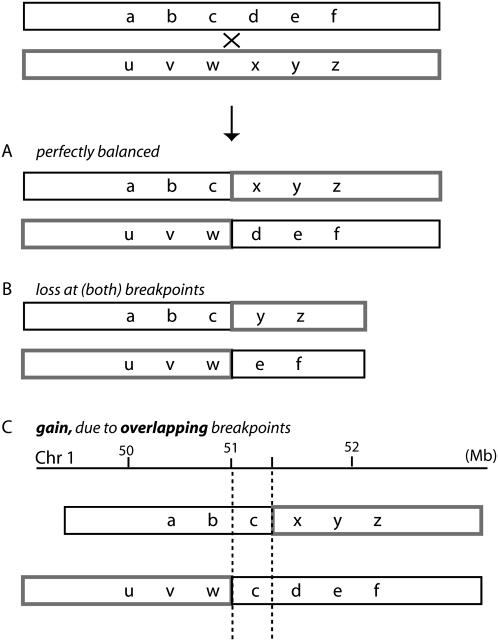
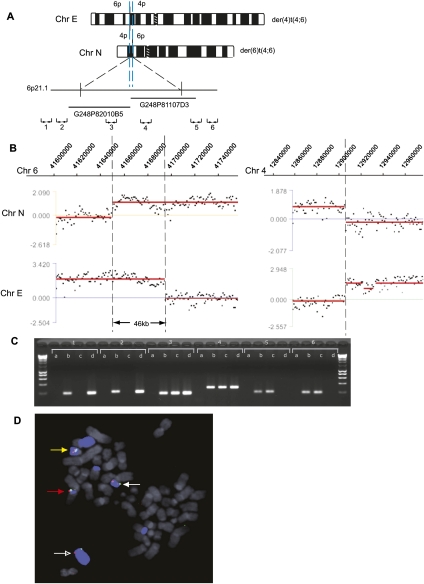
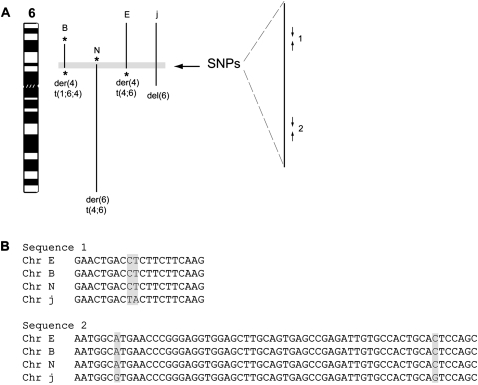
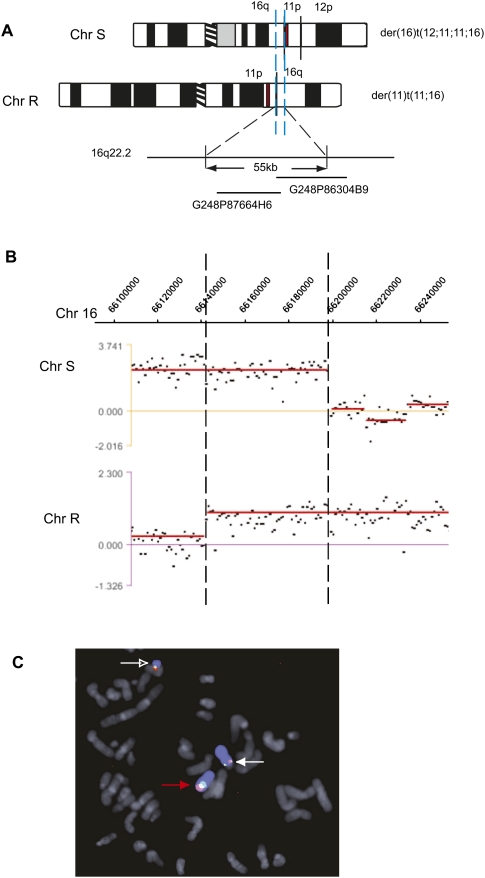
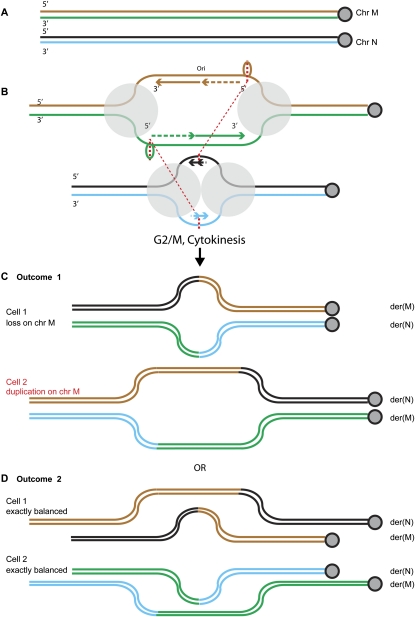
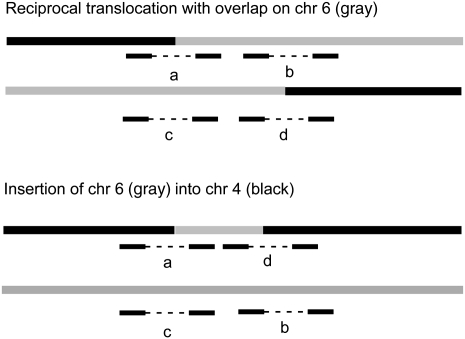
Reciprocal chromosome translocations are often not exactly reciprocal. Most familiar are deletions at the breakpoints, up to megabases in extent. We describe here the opposite phenomenon-duplication of tens or hundreds of kilobases at the breakpoint junction, so that the same sequence is present on both products of a translocation. When the products of the translocation are mapped on the genome, they overlap. We report several of these "overlapping-breakpoint" duplications in breast cancer cell lines HCC1187, HCC1806, and DU4475. These lines also had deletions and essentially balanced translocations. In HCC1187 and HCC1806, we identified five cases of duplication ranging between 46 kb and 200 kb, with the partner chromosome showing deletions between 29 bp and 31 Mb. DU4475 had a duplication of at least 200 kb. Breakpoints were mapped using array painting, i.e., hybridization of chromosomes isolated by flow cytometry to custom oligonucleotide microarrays. Duplications were verified by fluorescent in situ hybridization (FISH), PCR on isolated chromosomes, and cloning of breakpoints. We propose that these duplications are the counterpart of deletions and that they are produced at a replication bubble, comprising two replication forks with the duplicated sequence in between. Both copies of the duplicated sequence would go to one daughter cell, on different products of the translocation, while the other daughter cell would show deletion. These duplications may have been overlooked because they may be missed by FISH and array-CGH and may be interpreted as insertions by paired-end sequencing. Such duplications may therefore be quite frequent.
Figures
References
-
- Alsop AE, Taylor K, Zhang J, Gabra H, Paige AJ, Edwards PA 2008. Homozygous deletions may be markers of nearby heterozygous mutations: The complex deletion at FRA16D in the HCT116 colon cancer cell line removes exons of WWOX. Genes Chromosomes Cancer 47: 437–447 - PubMed
-
- Ciccone R, Giorda R, Gregato G, Guerrini R, Giglio S, Carrozzo R, Bonaglia MC, Priolo E, Lagana C, Tenconi R, et al. 2005. Reciprocal translocations: A trap for cytogenetists? Hum Genet 117: 571–582 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous