

Molecular basis of increased serum resistance among pulmonary isolates of non-typeable Haemophilus influenzae
- PMID: 21253576
- PMCID: PMC3017122
- DOI: 10.1371/journal.ppat.1001247
Molecular basis of increased serum resistance among pulmonary isolates of non-typeable Haemophilus influenzae
Abstract
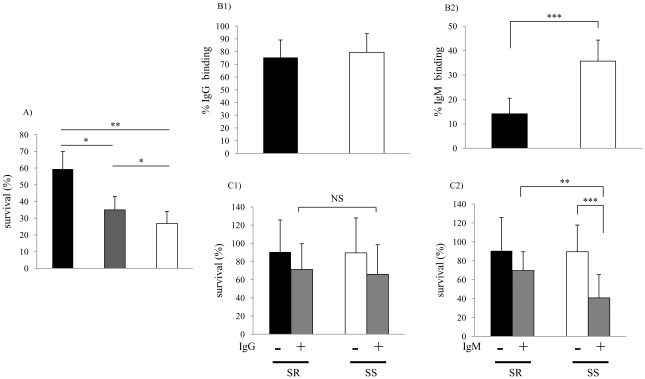
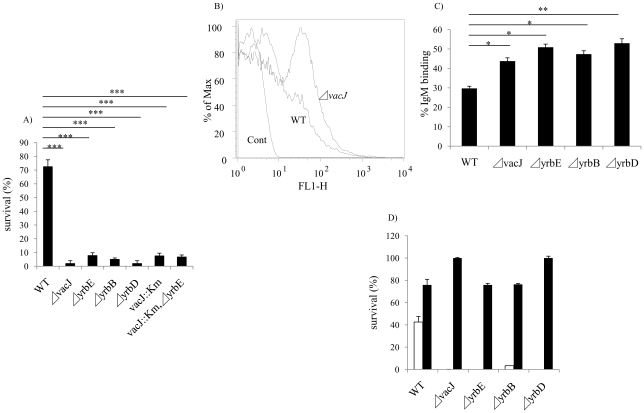
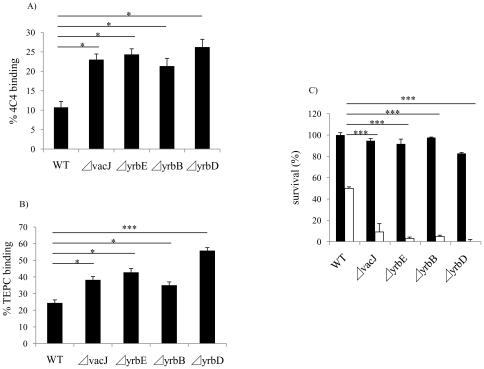
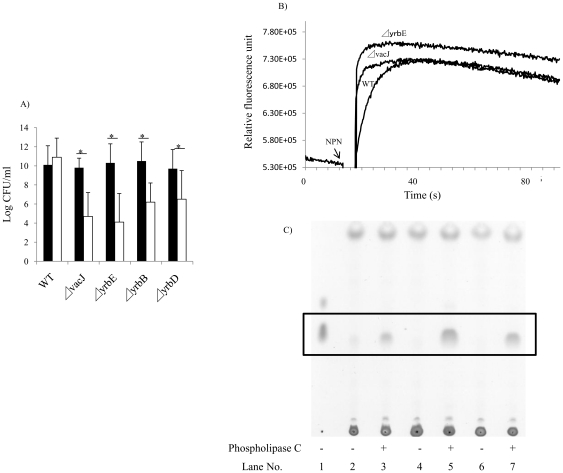
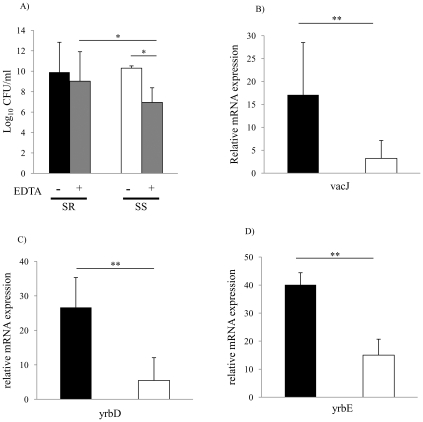
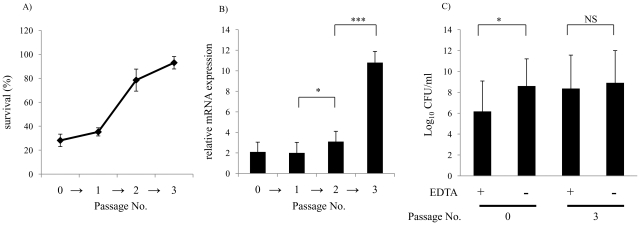
Non-typeable Haemophilus influenzae (NTHi), a common commensal of the human pharynx, is also an opportunistic pathogen if it becomes established in the lower respiratory tract (LRT). In comparison to colonizing isolates from the upper airway, LRT isolates, especially those associated with exacerbations of chronic obstructive pulmonary disease, have increased resistance to the complement- and antibody-dependent, bactericidal effect of serum. To define the molecular basis of this resistance, mutants constructed in a serum resistant strain using the mariner transposon were screened for loss of survival in normal human serum. The loci required for serum resistance contribute to the structure of the exposed surface of the bacterial outer membrane. These included loci involved in biosynthesis of the oligosaccharide component of lipooligosaccharide (LOS), and vacJ, which functions with an ABC transporter encoded by yrb genes in retrograde trafficking of phospholipids from the outer to inner leaflet of the cell envelope. Mutations in vacJ and yrb genes reduced the stability of the outer membrane and were associated with increased cell surface hyrophobicity and phospholipid content. Loss of serum resistance in vacJ and yrb mutants correlated with increased binding of natural immunoglobulin M in serum as well as anti-oligosaccharide mAbs. Expression of vacJ and the yrb genes was positively correlated with serum resistance among clinical isolates. Our findings suggest that NTHi adapts to inflammation encountered during infection of the LRT by modulation of its outer leaflet through increased expression of vacJ and yrb genes to minimize recognition by bactericidal anti-oligosaccharide antibodies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Murphy T. Respiratory infections caused by non-typeable Haemophilus influenzae. Curr Opin Infect Dis. 2003;16:129–134. - PubMed
-
- Turk DC. The pathogenicity of Haemophilus influenzae. J Med Microbiol. 1984;18:1–16. - PubMed
-
- Sethi S, Evans N, Grant BJ, Murphy TF. New strains of bacteria and exacerbations of chronic obstructive pulmonary disease. N Engl J Med. 2002;347:465–471. - PubMed
-
- Bandi V, Apicella MA, Mason E, Murphy TF, Siddiqi A, et al. Nontypeable Haemophilus influenzae in the lower respiratory tract of patients with chronic bronchitis. Am J Respir Crit Care Med. 2001;164:2114–2119. - PubMed
Publication types
MeSH terms
Grants and funding
- R01 AI044231/AI/NIAID NIH HHS/United States
- AI19641/AI/NIAID NIH HHS/United States
- R21 AI044231/AI/NIAID NIH HHS/United States
- R01 DC005840/DC/NIDCD NIH HHS/United States
- T32 AI060516/AI/NIAID NIH HHS/United States
- R37 AI038446/AI/NIAID NIH HHS/United States
- R01 HL083893/HL/NHLBI NIH HHS/United States
- R01 AI046512/AI/NIAID NIH HHS/United States
- R01 AI019641/AI/NIAID NIH HHS/United States
- R01 AI078538/AI/NIAID NIH HHS/United States
- AI44231/AI/NIAID NIH HHS/United States
- HL083893/HL/NHLBI NIH HHS/United States
- DC05840/DC/NIDCD NIH HHS/United States
- AI046512/AI/NIAID NIH HHS/United States
- AI38446/AI/NIAID NIH HHS/United States
- R01 AI038446/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
