

Fast oscillatory activity induced by kainate receptor activation in the rat basolateral amygdala in vitro
- PMID: 21255131
- PMCID: PMC3627304
- DOI: 10.1111/j.1460-9568.2010.07582.x
Fast oscillatory activity induced by kainate receptor activation in the rat basolateral amygdala in vitro
Abstract
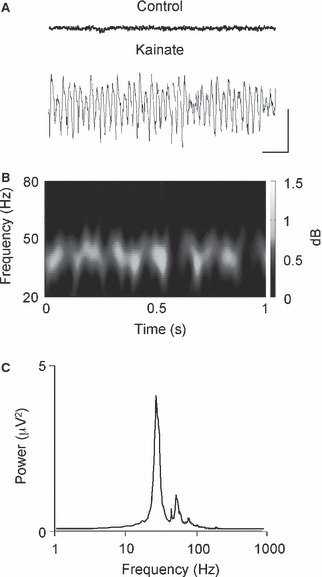
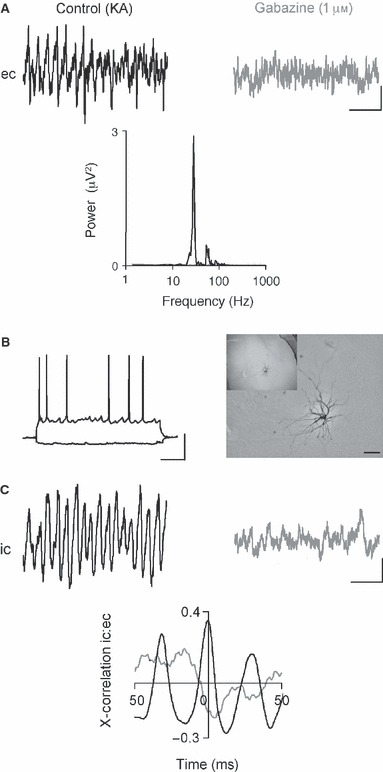
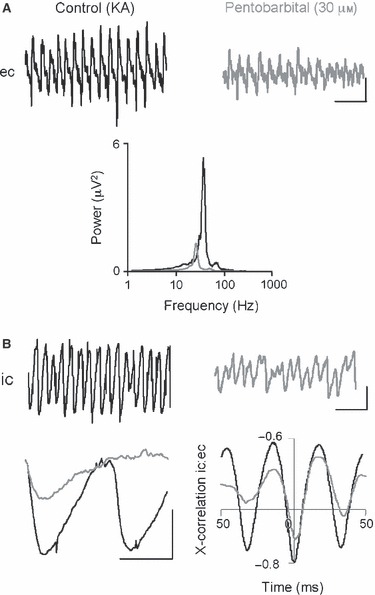
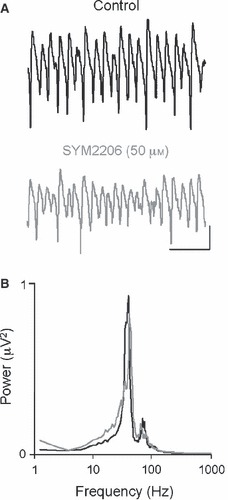
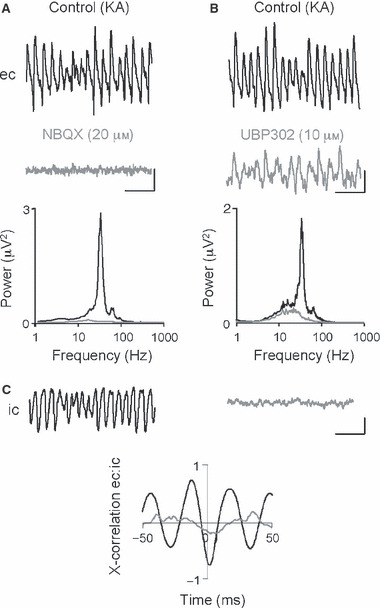
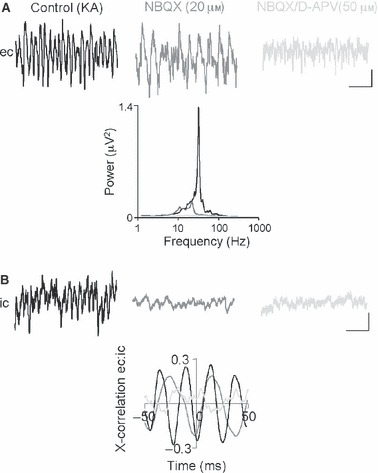
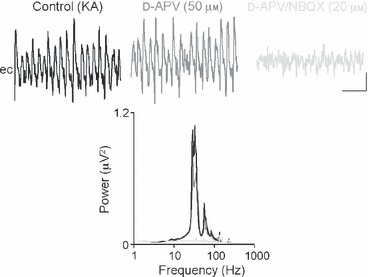
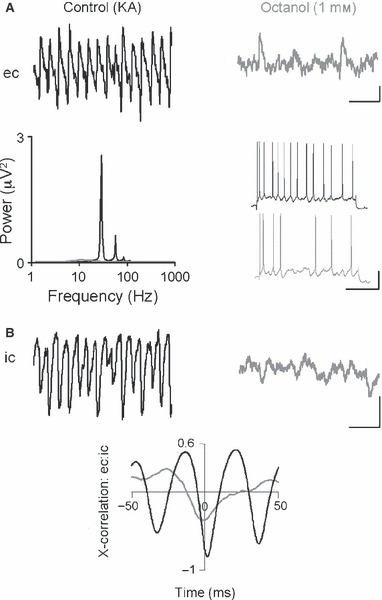
The basolateral amygdala (BLA) has a fundamental role in affective processing. In vivo studies have revealed rhythmic population activity of a similar type to that seen in the hippocampus and cortical areas during learning tasks. The amygdala contains densely interconnected networks of inhibitory interneurons similar to those responsible for fast network activity generation in the hippocampus and other cortical structures. Here we report that neuronal networks of the BLA in isolation generate persistent, gamma frequency (30-80 Hz) oscillations upon kainate receptor activation with kainic acid. We show that, like other cortical structures, BLA oscillations are completely dependent upon γ-aminobutyric acid (GABA)ergic inhibition. GABA(A) receptor blockade abolished all oscillations, and the activity was also sensitive to the barbiturate, pentobarbital. Blockade of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors or N-methyl-D-aspartate (NMDA) receptors had no significant effect on gamma activity. However, the GluR5-containing kainate receptor-specific antagonist (S)-1-(2-amino-2-carboxyethyl)-3-(2-carboxybenzyl) pyrimidine-2,4-dione (UBP302) abolished oscillations-evidence that glutamatergic receptor involvement is predominantly kainate receptor mediated. The mixed AMPA/kainate receptor antagonist 6-nitro-7-sulphamoylbenzo[f]quinoxalone-2,3-dione disodium (NBQX) abolished all oscillatory activity in 8/14 of slices tested. In the remaining slices, gamma frequency activity was abolished to reveal a low-amplitude, NMDA receptor-dependent, beta frequency (10-20 Hz) oscillation. Gamma oscillations are abolished by gap junction blockade. While these data show the BLA capable of generating gamma rhythms in common with other cortical areas studied to date, the network mechanisms appear to be different, suggesting a unique network structure underlies amygdala rhythmogenesis. Understanding how BLA networks produce synchronous activity is paramount to understanding how the BLA executes influence on important cognitive processes such as emotional learning.
© 2011 The Authors. European Journal of Neuroscience © 2011 Federation of European Neuroscience Societies and Blackwell Publishing Ltd.
Figures
References
-
- Bibbig A, Middleton S, Racca C, Gillies MJ, Garner H, Lebeau FE, Davies CH, Whittington MA. Beta rhythms (15-20 Hz) generated by nonreciprocal communication in hippocampus. J. Neurophysiol. 2007;97:2812–2823. - PubMed
-
- Buzsaki G. Two-stage model of memory trace formation: a role for “noisy” brain states. Neuroscience. 1989;31:551–570. - PubMed
-
- Cahill L. Neurobiological mechanisms of emotionally influenced, long-term memory. Prog. Brain Res. 2000;126:29–37. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
