

Antibody-mediated neutralization of flaviviruses: a reductionist view
- PMID: 21255816
- PMCID: PMC3100196
- DOI: 10.1016/j.virol.2010.12.020
Antibody-mediated neutralization of flaviviruses: a reductionist view
Abstract
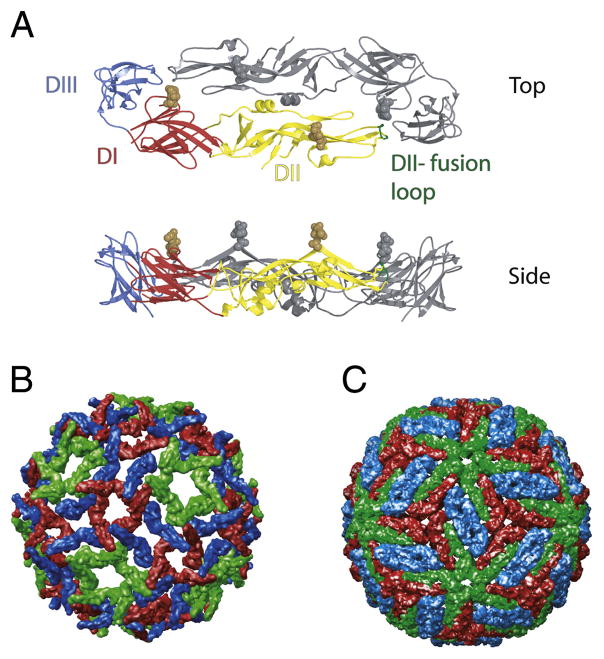
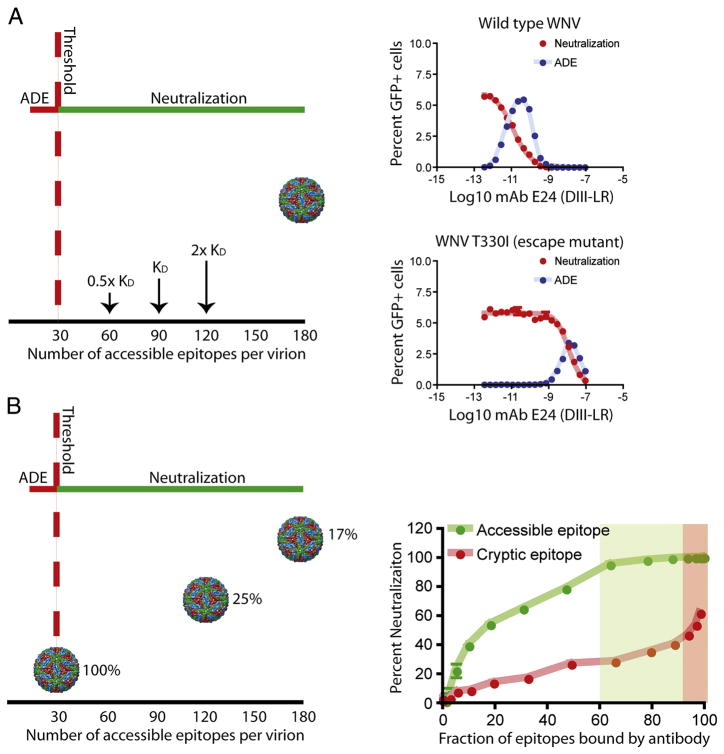
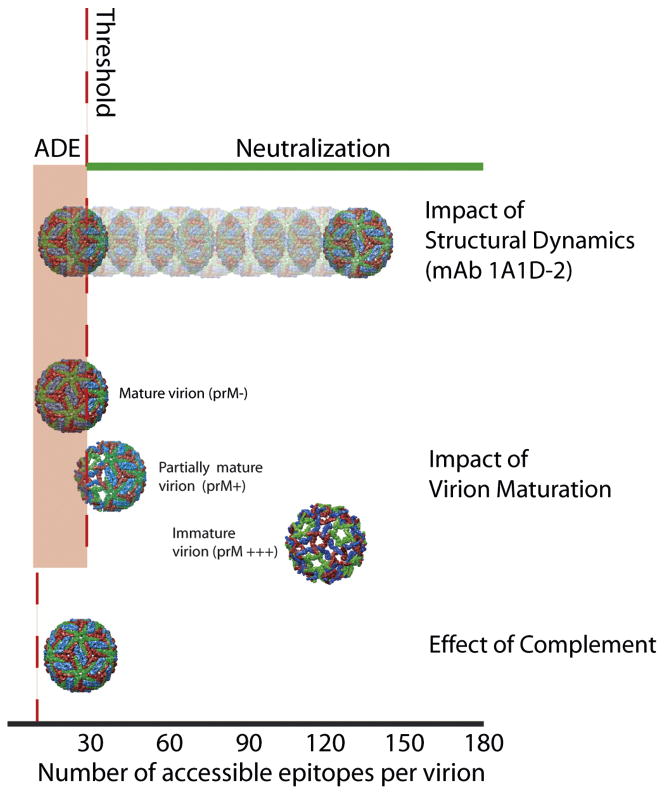
Flaviviruses are a group of ~70 small RNA viruses responsible for significant morbidity and mortality across the globe. Efforts to develop effective vaccines for several clinically important flaviviruses are underway. Antibodies are a significant component of the host's protective response against flavivirus infection with the potential to contribute to immunity via several distinct mechanisms, including an ability to directly neutralize virus infection. Conversely, virus-reactive antibodies have been implicated in the increased risk of severe clinical manifestations following secondary dengue virus infection. In this review, we will discuss recent progress toward understanding the molecular basis of antibody-mediated neutralization of flaviviruses. Neutralization requires engagement of the virion with a stoichiometry that exceeds a required threshold. From this perspective, we will discuss viral and host factors that impact the number of antibody molecules bound to the virus particle and significantly modulate the potency of neutralizing antibodies.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Aaskov JG, Williams L, Fletcher J, Hay R. Failure of a dengue 1 subunit vaccine to protect mice against a lethal dengue virus infection. Am J Trop Med Hyg. 1988;39(5):511–8. - PubMed
-
- Barrett AD, Teuwen DE. Yellow fever vaccine - how does it work and why do rare cases of serious adverse events take place? Curr Opin Immunol. 2009;21(3):308–13. - PubMed
-
- Beltramello M, Williams KL, Simmons CP, Macagno A, Simonelli L, Quyen NT, Sukupolvi-Petty S, Navarro-Sanchez E, Young PR, de Silva AM, Rey FA, Varani L, Whitehead SS, Diamond MS, Harris E, Lanzavecchia A, Sallusto F. The human immune response to Dengue virus is dominated by highly cross-reactive antibodies endowed with neutralizing and enhancing activity. Cell Host Microbe. 2010;8(3):271–83. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
