

The Ndc80 loop region facilitates formation of kinetochore attachment to the dynamic microtubule plus end
- PMID: 21256019
- PMCID: PMC3052438
- DOI: 10.1016/j.cub.2010.12.050
The Ndc80 loop region facilitates formation of kinetochore attachment to the dynamic microtubule plus end
Abstract
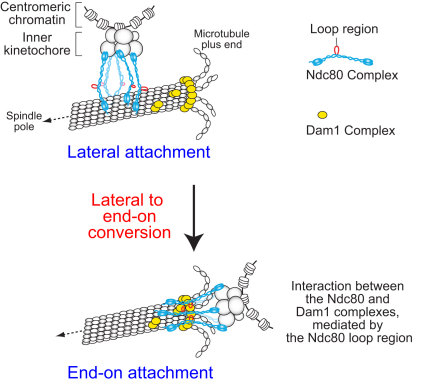
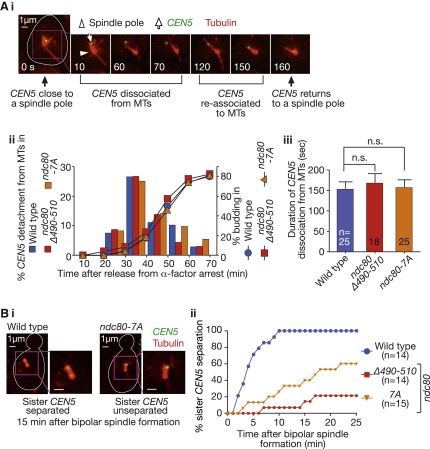
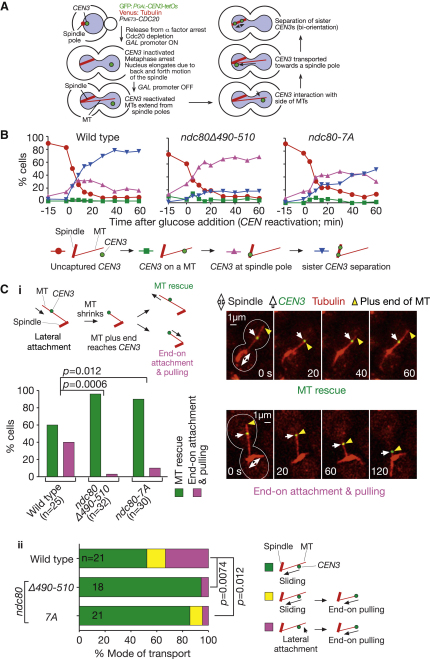
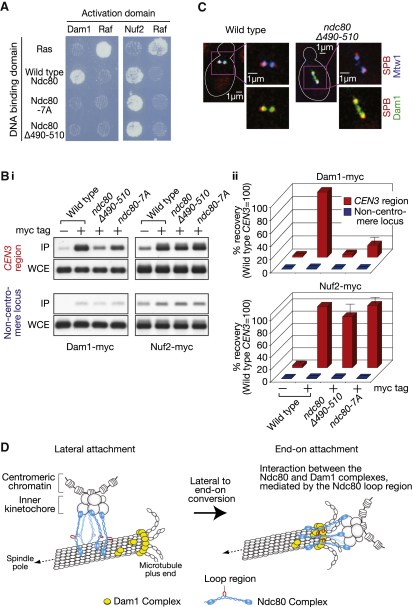
Proper chromosome segregation in mitosis relies on correct kinetochore-microtubule (KT-MT) interactions. The KT initially interacts with the lateral surface of a single MT (lateral attachment) extending from a spindle pole and is subsequently anchored at the plus end of the MT (end-on attachment). The conversion from lateral to end-on attachment is crucial because end-on attachment is more robust and thought to be necessary to sustain KT-MT attachment when tension is applied across sister KTs upon their biorientation. The mechanism for this conversion is still elusive. The Ndc80 complex is an essential component of the KT-MT interface, and here we studied a role of the Ndc80 loop region, a distinct motif looping out from the coiled-coil shaft of the complex, in Saccharomyces cerevisiae. With deletions or mutations of the loop region, the lateral KT-MT attachment occurred normally; however, subsequent conversion to end-on attachment was defective, leading to failure in sister KT biorientation. The Ndc80 loop region was required for Ndc80-Dam1 interaction and KT loading of the Dam1 complex, which in turn supported KT tethering to the dynamic MT plus end. The Ndc80 loop region, therefore, has an important role in the conversion from lateral to end-on attachment, a crucial maturation step of KT-MT interaction.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures

Comment in
-
Chromosome segregation: keeping kinetochores in the loop.Curr Biol. 2011 Feb 8;21(3):R110-2. doi: 10.1016/j.cub.2010.12.030. Curr Biol. 2011. PMID: 21300272
References
-
- Grishchuk E.L., Molodtsov M.I., Ataullakhanov F.I., McIntosh J.R. Force production by disassembling microtubules. Nature. 2005;438:384–388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
