

Ndc80 internal loop interacts with Dis1/TOG to ensure proper kinetochore-spindle attachment in fission yeast
- PMID: 21256022
- PMCID: PMC3049873
- DOI: 10.1016/j.cub.2010.12.048
Ndc80 internal loop interacts with Dis1/TOG to ensure proper kinetochore-spindle attachment in fission yeast
Abstract
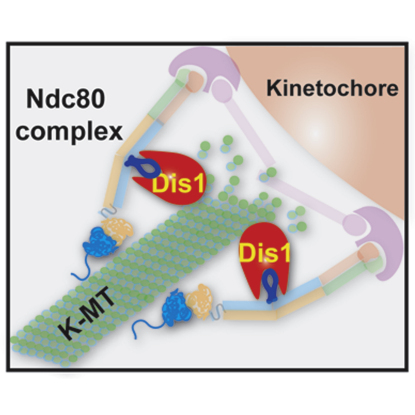
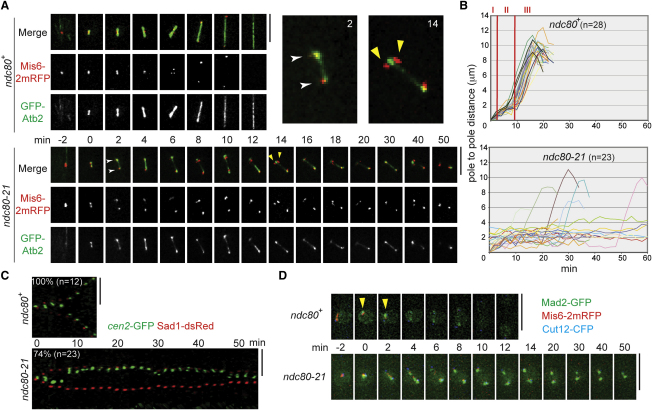
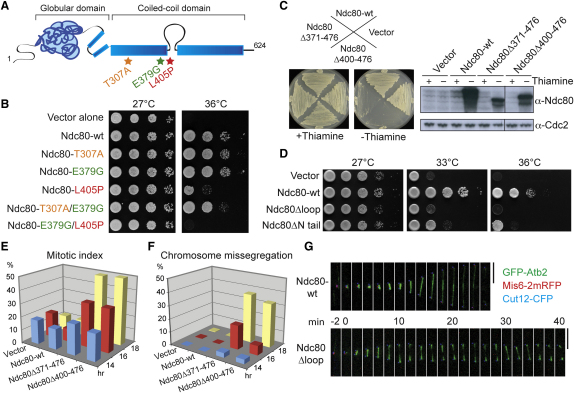
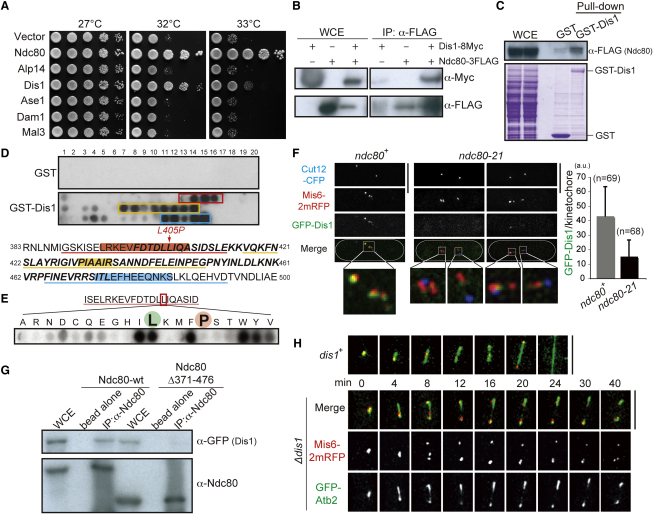
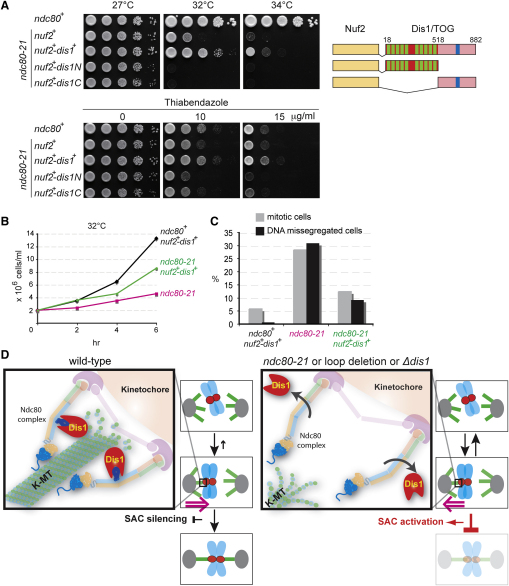
The Ndc80 complex, a conserved outer kinetochore complex, comprising four components (Ndc80/Hec1, Nuf2, Spc24, and Spc25), constitutes one of the core microtubule-binding sites within the kinetochore. Despite this knowledge, molecular mechanisms by which this complex contributes to establishment of correct bipolar attachment of the kinetochore to the spindle microtubule remain largely elusive. Here we show that the conserved internal loop of fission yeast Ndc80 directly binds the Dis1/TOG microtubule-associated protein, thereby coupling spindle microtubule dynamics with kinetochore capture. Ndc80 loop mutant proteins fail to recruit Dis1 to kinetochores, imposing unstable attachment and frequent spindle collapse. In these mutants, mitotic progression is halted attributable to spindle assembly checkpoint activation, and chromosomes remain in the vicinity of the spindle poles without congression. dis1 deletion precisely phenocopies the loop mutants. Intriguingly, forced targeting of Dis1 to the Ndc80 complex rescues loop mutant's defects. We propose that Ndc80 comprises two microtubule-interacting interfaces: the N-terminal region directly binds the microtubule lattice, while the internal loop interacts with the plus end of microtubules via Dis1/TOG. Therefore, our results provide a crucial insight into how the Ndc80 complex establishes stable bipolar attachment to the spindle microtubule.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Chromosome segregation: keeping kinetochores in the loop.Curr Biol. 2011 Feb 8;21(3):R110-2. doi: 10.1016/j.cub.2010.12.030. Curr Biol. 2011. PMID: 21300272
References
-
- Cheeseman I.M., Chappie J.S., Wilson-Kubalek E.M., Desai A. The conserved KMN network constitutes the core microtubule-binding site of the kinetochore. Cell. 2006;127:983–997. - PubMed
-
- Wei R.R., Al-Bassam J., Harrison S.C. The Ndc80/HEC1 complex is a contact point for kinetochore-microtubule attachment. Nat. Struct. Mol. Biol. 2007;14:54–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
