

doi: 10.1038/nsmb.1976.
Epub 2011 Jan 23.
Dynamic local unfolding in the serpin α-1 antitrypsin provides a mechanism for loop insertion and polymerization
Affiliations
- PMID: 21258324
- PMCID: PMC3074950
- DOI: 10.1038/nsmb.1976
Item in Clipboard
Dynamic local unfolding in the serpin α-1 antitrypsin provides a mechanism for loop insertion and polymerization
Nat Struct Mol Biol.
2011 Feb.
Abstract
The conformational plasticity of serine protease inhibitors (serpins) underlies both their activities as protease inhibitors and their susceptibility to pathogenic misfolding and aggregation. Here, we structurally characterize a sheet-opened state of the serpin α-1 antitrypsin (α₁AT) and show how local unfolding allows functionally essential strand insertion. Mutations in α₁AT that cause polymerization-induced serpinopathies map to the labile region, suggesting that the evolution of serpin function required sampling of high risk conformations on a dynamic energy landscape.
Figures
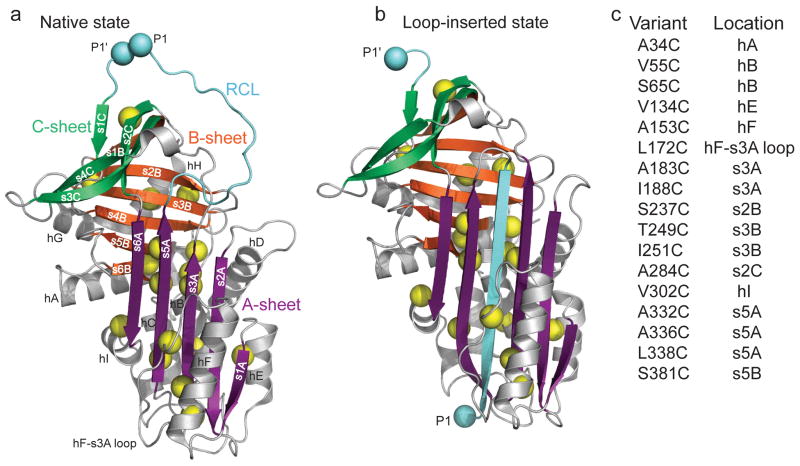
Design of cysteine mutations to probe low denaturant-induced strand opening in the serpin α1AT. Comparison of structures of (a) native (PDB 1QLP) and (b) cleaved ‘loop-inserted’ (PDB 1EZX) states of α1AT. The two light blue spheres, labeled P1 and P1′, correspond to the protease cleavage site in the RCL. Sites mutated to cysteine are indicated by yellow spheres centered on the Cβ. (c) Positions of cysteine substitutions and their secondary structural context. Structures in this and other figures were prepared using PyMol (http://www.pymol.org ).
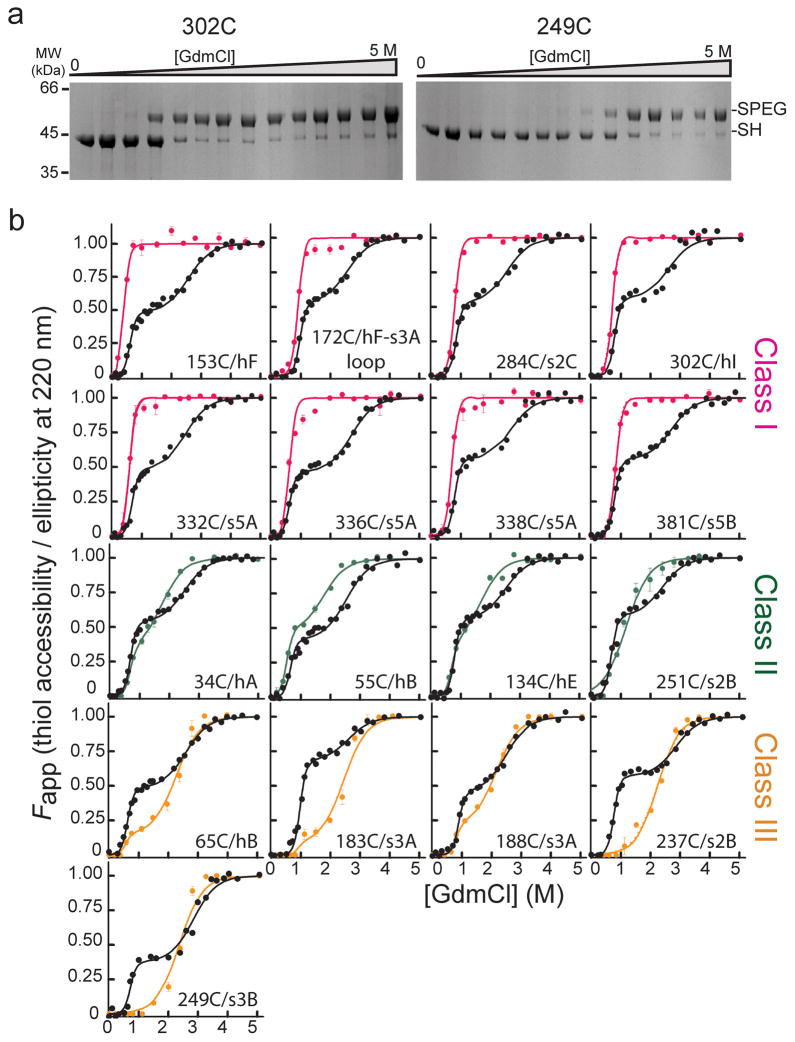
Accessibility of α1AT single cysteine variants as a function of denaturant and comparison to unfolding monitored by CD. (a) Coomassie-stained SDS-PAGE results for the PEGylation of two representative single cysteine variants as a function of GdmCl concentration. Bands corresponding to the free and PEG-modified thiol are indicated by ‘-SH’ and ‘-SPEG’, respectively. (b) Comparison of the fractional accessibility of each cysteine to PEGylation (colored circles) with the unfolding of the variant α1AT monitored by CD (black circles). The solid lines through the data are the fit to a three-state or a two-state protein unfolding model as described in data analyses (see Supplementary Tables 2 and 3 for fit data). Data are grouped and colored according to the behavior of the specific cysteine site as a function of denaturant, i.e., Class I sites, which become fully accessible at low denaturant, are shown in pink, Class II sites, which become nearly 50% accessible at low denaturant and fully accessible as the protein globally unfolds, in green, and Class III sites, which are inaccessible or only slightly accessible over the first unfolding transition and then become fully accessible when the protein globally unfolds, in orange.
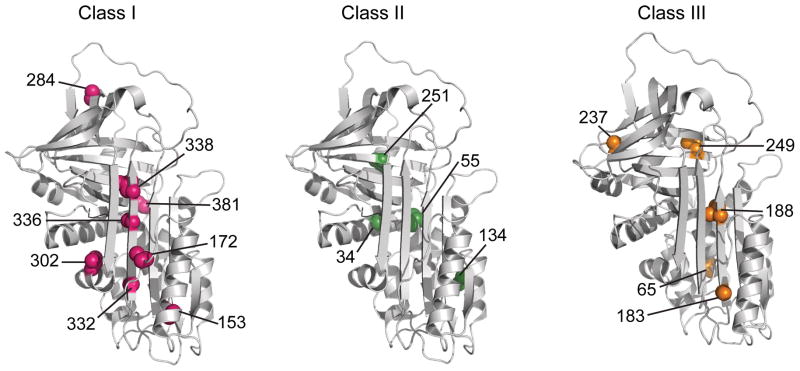
Structural context of the cysteines belonging to the three classes with varying extents of PEGylation. Class I (pink), II (green), and III (orange) exhibit 100%, ~50% and <20% PEGylation, respectively, in the low GdmCl intermediate state of α1AT. The seventeen residues investigated in the current study are shown on the native structure of α1AT (PDB ID 1QLP).
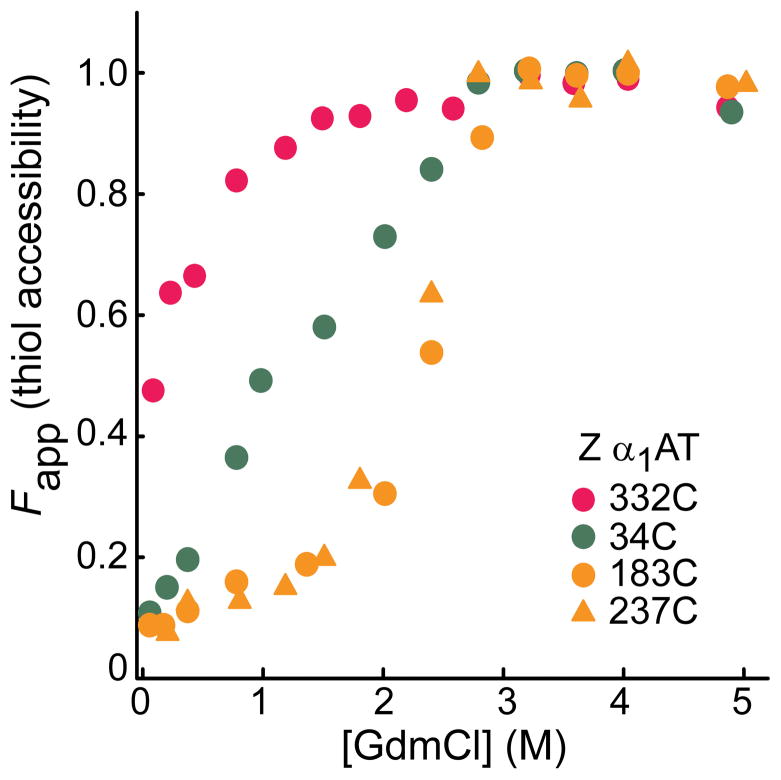
Solvent accessibility of single cysteines in the Z variant of α1AT. The intermediate state of the protein was formed from the denatured state, i.e., upon dilution from high (5M GdmCl) denaturant. Bands of the PEG-modified (-SPEG) and unmodified (-SH) protein in Supplementary Fig. 4a were quantitated to obtain fraction thiol accessibility. Note that refolding of the Z variant yields a state that is not identical to native, and that this accounts for the partial exposure of 332C at the lowest GdmCl concentration.
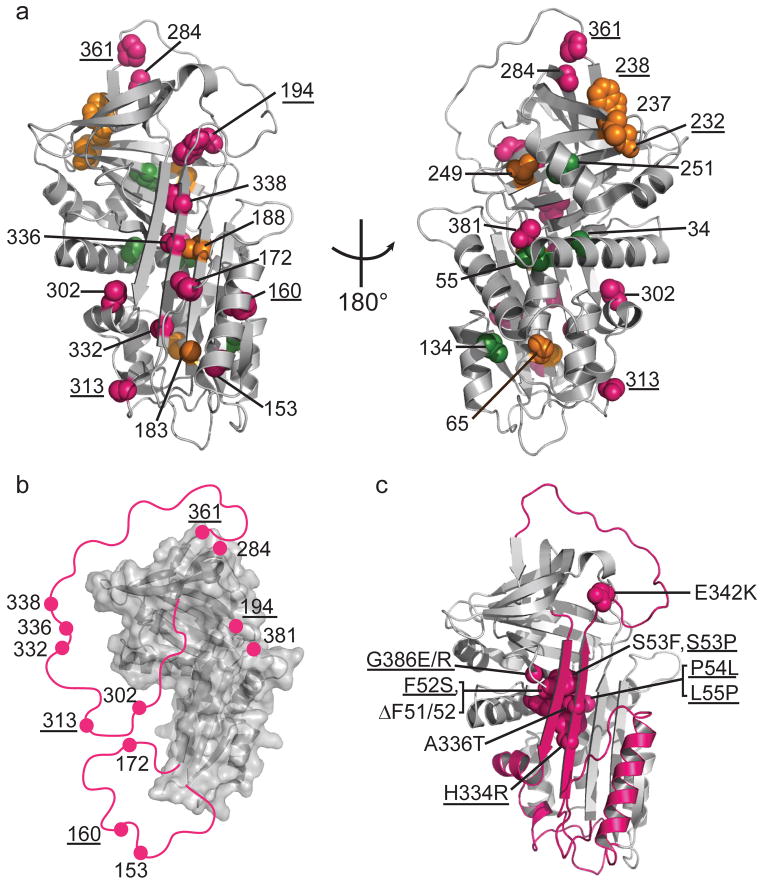
Local unfolding leads to the sheet-opened intermediate of α1AT. (a) Cysteine positions are shown on the structure of native α1AT (PDB 1QLP) and colored by their accessibility behavior with pink, green and orange representing Class I, Class II, and III, respectively (see Fig. 2 and text for description of classes). Results for an additional six residue positions from previous work,, are indicated with appropriate coloring on the structure as well (labels are underlined) using the same color scheme. (b) The stable structural elements of the sheet-opened intermediate as inferred from cysteine accessibility data. In this structure, regions found to be accessible in the sheet-opened intermediate of α1AT are removed and indicated schematically by a pink line connecting to the remaining structure. (c) The five naturally occurring pathogenic point mutations in α1AT that produce full-length, polymerogenic protein and cause liver damage, along with similar polymerogenic mutations reported for other serpins (underlined labels) are represented in pink spacefill on the native structure of α1AT. The backbone of the identified labile region is in pink.
Similar articles
-
Defining the mechanism of polymerization in the serpinopathies.Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17146-51. doi: 10.1073/pnas.1004785107. Epub 2010 Sep 20. Proc Natl Acad Sci U S A. 2010. PMID: 20855577 Free PMC article.
-
The structural basis of serpin polymerization studied by hydrogen/deuterium exchange and mass spectrometry.J Biol Chem. 2008 Nov 7;283(45):30804-11. doi: 10.1074/jbc.M804048200. Epub 2008 Sep 15. J Biol Chem. 2008. PMID: 18794298 Free PMC article.
-
Probing the local conformational change of alpha1-antitrypsin.Protein Sci. 2007 Sep;16(9):1842-50. doi: 10.1110/ps.072911607. Epub 2007 Jul 27. Protein Sci. 2007. PMID: 17660256 Free PMC article.
-
Conformational changes in serpins and the mechanism of alpha 1-antitrypsin deficiency.Am J Respir Crit Care Med. 1994 Dec;150(6 Pt 2):S171-5. doi: 10.1164/ajrccm/150.6_Pt_2.S171. Am J Respir Crit Care Med. 1994. PMID: 7952655 Review.
-
New insights into the structural basis of alpha 1-antitrypsin deficiency.QJM. 1996 Nov;89(11):807-12. doi: 10.1093/qjmed/89.11.807. QJM. 1996. PMID: 8977959 Review.
Cited by
-
How the serpin α1-proteinase inhibitor folds.J Biol Chem. 2012 Apr 6;287(15):12425-32. doi: 10.1074/jbc.M111.315465. Epub 2012 Feb 13. J Biol Chem. 2012. PMID: 22334651 Free PMC article.
-
Collapse of a long axis: single-molecule Förster resonance energy transfer and serpin equilibrium unfolding.Biochemistry. 2014 May 13;53(18):2903-14. doi: 10.1021/bi401622n. Epub 2014 May 1. Biochemistry. 2014. PMID: 24749911 Free PMC article.
-
The Z mutation alters the global structural dynamics of α1-antitrypsin.PLoS One. 2014 Sep 2;9(9):e102617. doi: 10.1371/journal.pone.0102617. eCollection 2014. PLoS One. 2014. PMID: 25181470 Free PMC article.
-
Challenges and Prospects for Alpha-1 Antitrypsin Deficiency Gene Therapy.Hum Gene Ther. 2015 Nov;26(11):709-18. doi: 10.1089/hum.2015.044. Epub 2015 Sep 29. Hum Gene Ther. 2015. PMID: 26413996 Free PMC article. Review.
-
ER chaperones use a protein folding and quality control glyco-code.Mol Cell. 2023 Dec 21;83(24):4524-4537.e5. doi: 10.1016/j.molcel.2023.11.006. Epub 2023 Dec 4. Mol Cell. 2023. PMID: 38052210 Free PMC article.
References
-
- Gettins PG. Serpin structure, mechanism, and function. Chem Rev. 2002;102:4751–4804. - PubMed
-
- Huntington JA, Read RJ, Carrell RW. Structure of a serpin-protease complex shows inhibition by deformation. Nature. 2000;407:923–926. - PubMed
-
- Dupont DM, et al. Biochemical properties of plasminogen activator inhibitor-1. Front Biosci. 2009;14:1337–1361. - PubMed
-
- Mushunje A, Evans G, Brennan SO, Carrell RW, Zhou A. Latent antithrombin and its detection, formation and turnover in the circulation. J Thromb Haemost. 2004;2:2170–2177. - PubMed
-
- Kaslik G, et al. Effects of serpin binding on the target proteinase: global stabilization, localized increased structural flexibility, and conserved hydrogen bonding at the active site. Biochemistry. 1997;36:5455–5464. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
