

Chromatin accessibility pre-determines glucocorticoid receptor binding patterns
- PMID: 21258342
- PMCID: PMC6386452
- DOI: 10.1038/ng.759
Chromatin accessibility pre-determines glucocorticoid receptor binding patterns
Abstract
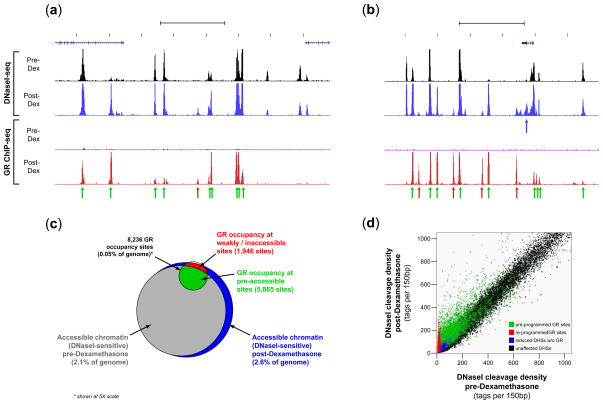
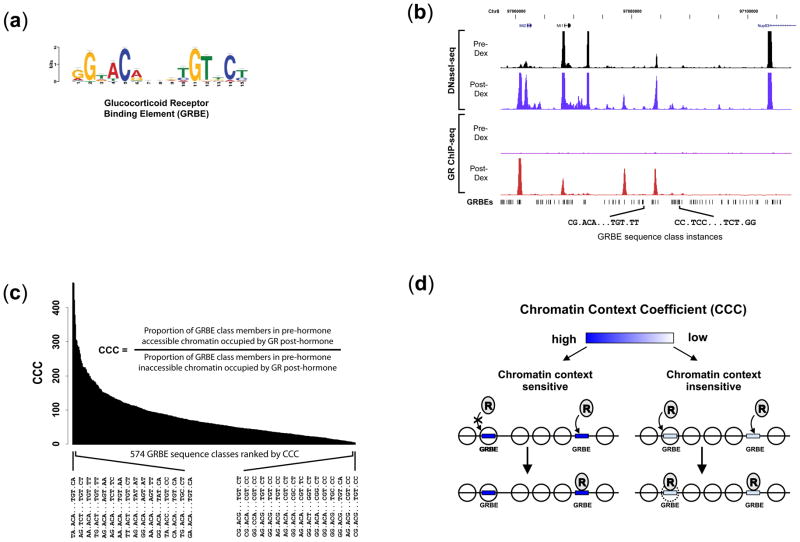
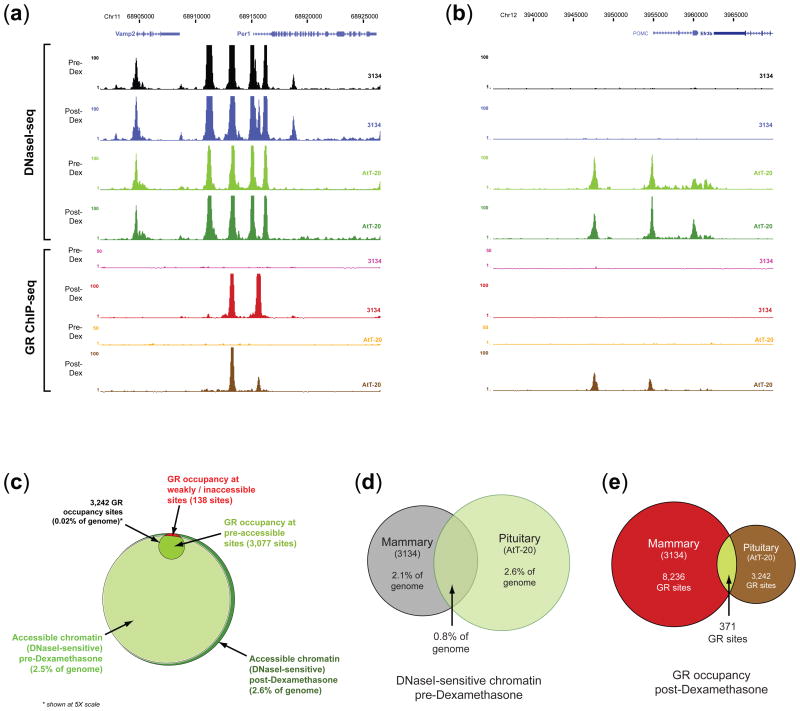
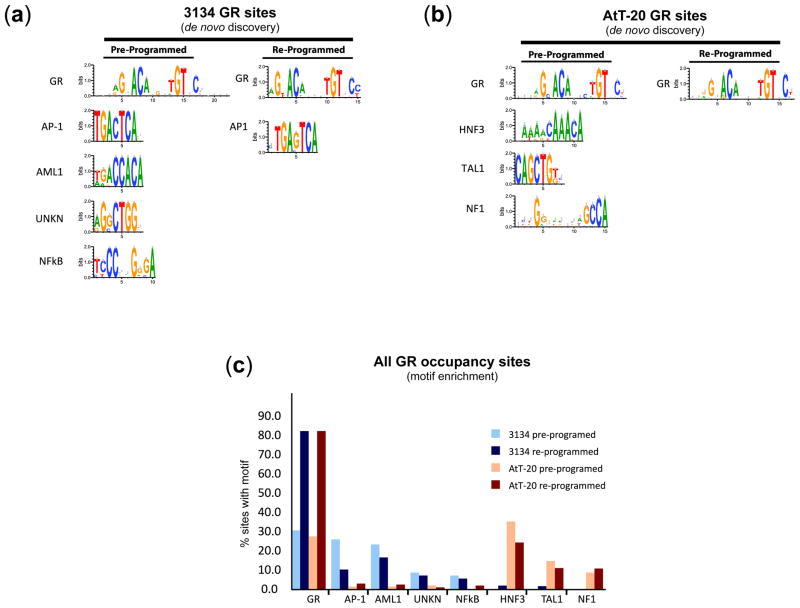
Development, differentiation and response to environmental stimuli are characterized by sequential changes in cellular state initiated by the de novo binding of regulated transcriptional factors to their cognate genomic sites. The mechanism whereby a given regulatory factor selects a limited number of in vivo targets from a myriad of potential genomic binding sites is undetermined. Here we show that up to 95% of de novo genomic binding by the glucocorticoid receptor, a paradigmatic ligand-activated transcription factor, is targeted to preexisting foci of accessible chromatin. Factor binding invariably potentiates chromatin accessibility. Cell-selective glucocorticoid receptor occupancy patterns appear to be comprehensively predetermined by cell-specific differences in baseline chromatin accessibility patterns, with secondary contributions from local sequence features. The results define a framework for understanding regulatory factor-genome interactions and provide a molecular basis for the tissue selectivity of steroid pharmaceuticals and other agents that intersect the living genome.
Figures
References
-
- Britten RJ, Davidson EH. Gene regulation for higher cells: a theory. Science. 1969;165:349–357. - PubMed
-
- McKenna NJ, O'Malley BW. Combinatorial control of gene expression by nuclear receptors and coregulators. Cell. 2002;108:465–474. - PubMed
-
- Okita K, Ichisaka T, Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007;448:313–317. - PubMed
-
- Felsenfeld G, Groudine M. Controlling the double helix. Nature. 2003;421:448–453. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
