

Tracing explosives in soil with transcriptional regulators of Pseudomonas putida evolved for responding to nitrotoluenes
- PMID: 21261843
- PMCID: PMC3815885
- DOI: 10.1111/j.1751-7915.2008.00027.x
Tracing explosives in soil with transcriptional regulators of Pseudomonas putida evolved for responding to nitrotoluenes
Abstract
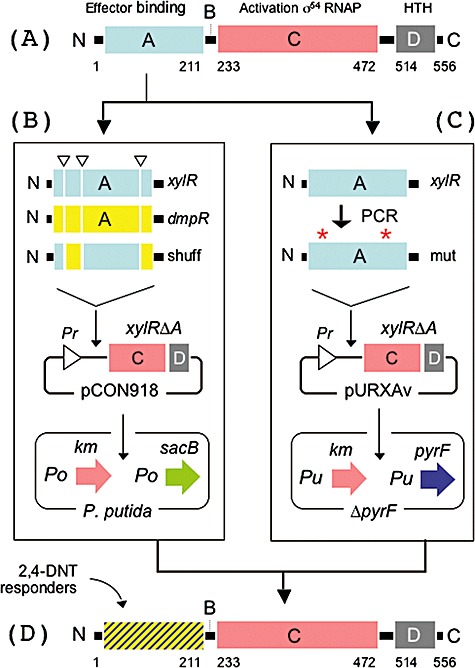
Although different biological approaches for detection of anti-personnel mines and other unexploded ordnance (UXO) have been entertained, none of them has been rigorously documented thus far in the scientific literature. The industrial 2,4,6 trinitrotoluene (TNT) habitually employed in the manufacturing of mines is at all times tainted with a small but significant proportion of the more volatile 2,4 dinitrotoluene (2,4 DNT) and other nitroaromatic compounds. By using mutation-prone PCR and DNA sequence shuffling we have evolved in vitro and selected in vivo variants of the effector recognition domain of the toluene-responsive XylR regulator of the soil bacterium Pseudomonas putida that responds to mono-, bi- and trinitro substituted toluenes. Re-introduction of such variants in P. putida settled the transcriptional activity of the cognate promoters (Po and Pu) as a function of the presence of nitrotoluenes in the medium. When strains bearing transcriptional fusions to reporters with an optical output (luxAB, GFP) were spread on soil spotted with nitrotoluenes, the signal triggered by promoter activation allowed localization of the target compounds on the soil surface. Our data provide a proof of concept that non-natural transcription factors evolved to respond to nitroaromatics can be engineered in soil bacteria and inoculated on a target site to pinpoint the presence of explosives. This approach thus opens new ways to tackle this gigantic humanitarian problem.
© 2008 The Authors.
Figures





References
-
- Altamirano M., Garcia‐Villada L., Agrelo M., Sanchez‐Martin L., Martin‐Otero L., Flores‐Moya A. A novel approach to improve specificity of algal biosensors using wild‐type and resistant mutants: an application to detect TNT. Biosens Bioelectron. 2004;19:1319–1323. et al. - PubMed
-
- Blatny J.M., Brautaset T., Winther‐Larsen H.C., Karunakaran P., Valla S. Improved broad‐host‐range RK2 vectors useful for high and low regulated gene expression levels in gram‐negative bacteria. Plasmid. 1997;38:35–51. - PubMed
-
- Bromenshenk J., Henderson C.B., Smith G.C. Biological systems (paper II) In: MacDonald J., Lockwood J.R., McFee J., Altshuler T., Broach T., Carin L., editors. RAND Corporation; 2003. pp. 273–283. , and . In Alternatives for Landmine Detection. et al. (eds). Santa Monica, CA, USA: , pp.
-
- Bruschini C., Gros B. A survey of current sensor technology research for the detection of landmines. Proceedings of the International Workshop on Sustainable Humanitarian Demining, Zagreb, Croatia. 1997;SusDem‐97:6.18–16.27.
-
- Bundy J.G., Durham D.G., Paton G.I., Campbell C.D. Investigating the specificity of regulators of degradation of hydrocarbons and hydrocarbon‐based compounds using structure–activity relationships. Biodegradation. 2000;11:37–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
