

Amyloid beta dimers/trimers potently induce cofilin-actin rods that are inhibited by maintaining cofilin-phosphorylation
- PMID: 21261978
- PMCID: PMC3037337
- DOI: 10.1186/1750-1326-6-10
Amyloid beta dimers/trimers potently induce cofilin-actin rods that are inhibited by maintaining cofilin-phosphorylation
Abstract
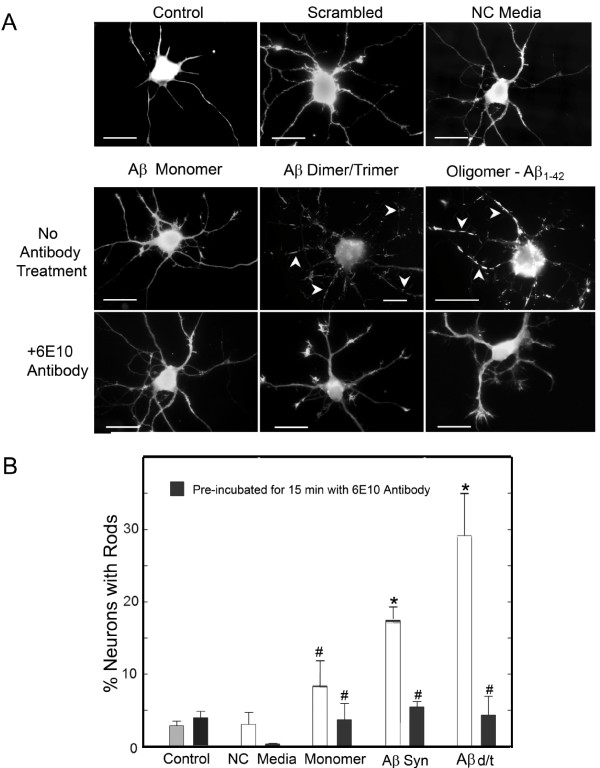
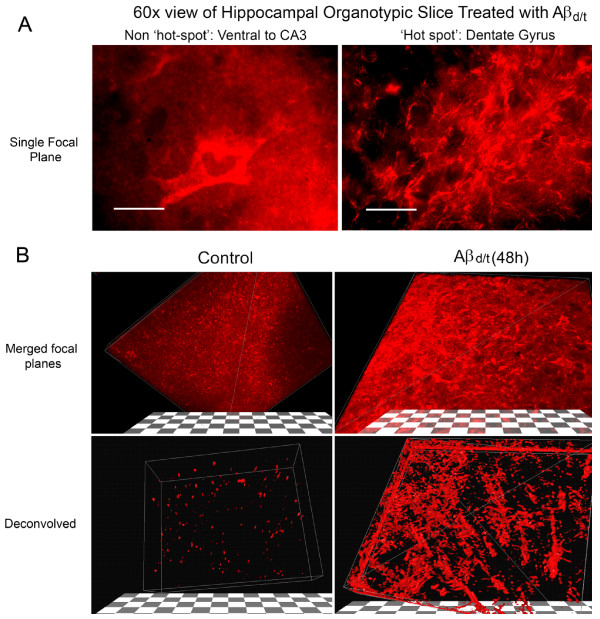
Background: Previously we reported 1 μM synthetic human amyloid beta1-42 oligomers induced cofilin dephosphorylation (activation) and formation of cofilin-actin rods within rat hippocampal neurons primarily localized to the dentate gyrus.
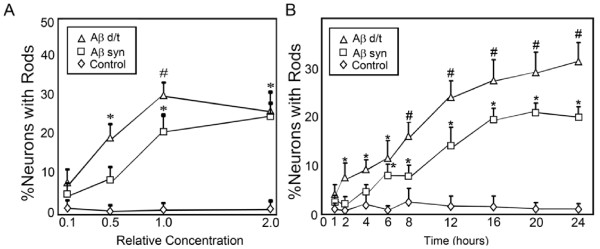
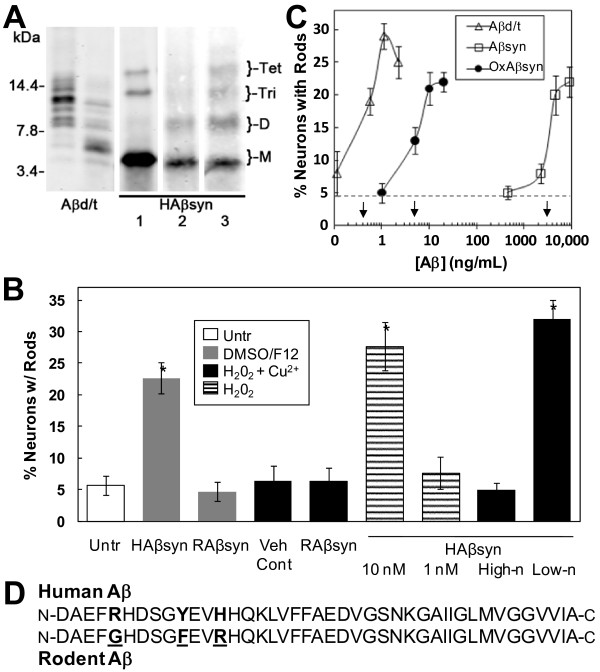
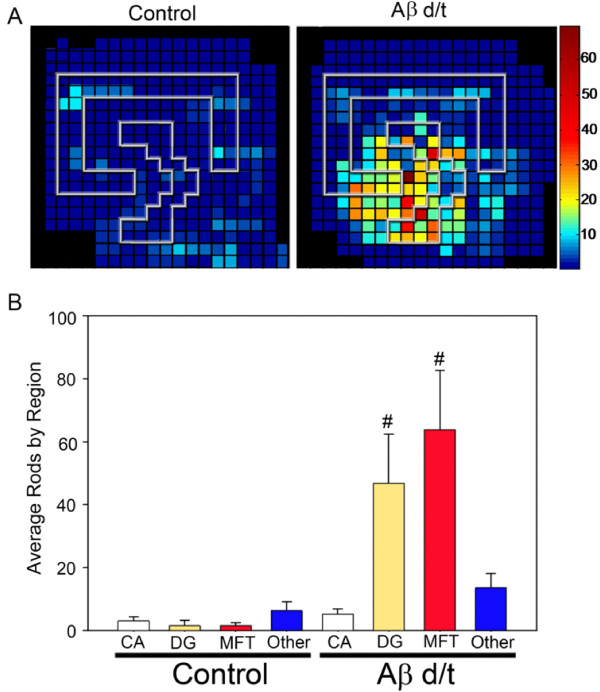
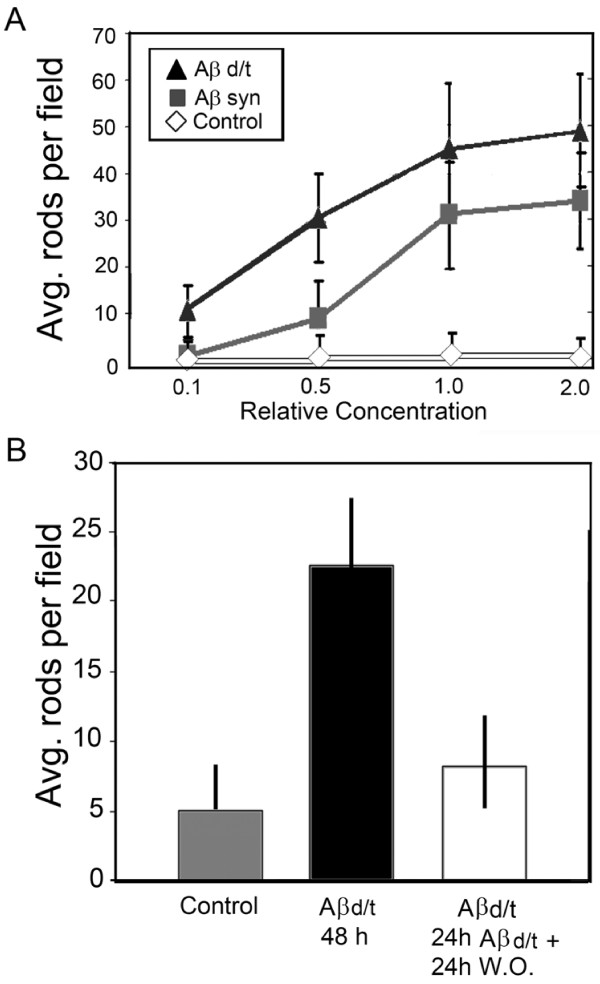
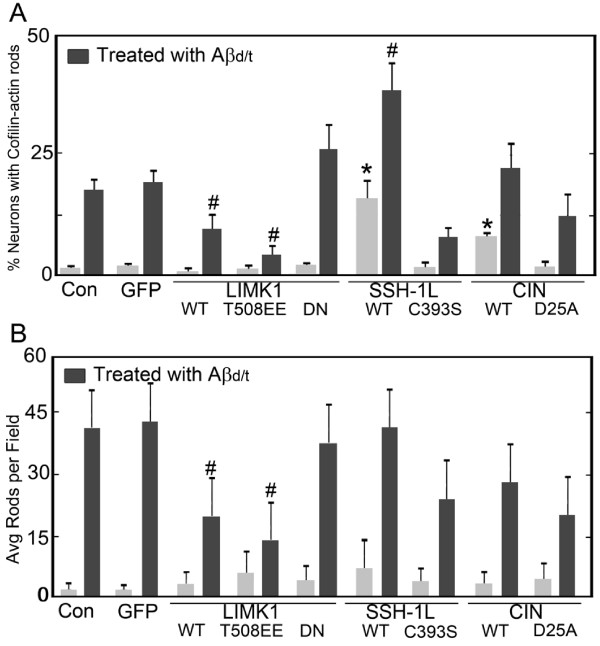
Results: Here we demonstrate that a gel filtration fraction of 7PA2 cell-secreted SDS-stable human Aβ dimers and trimers (Aβd/t) induces maximal neuronal rod response at ~250 pM. This is 4,000-fold more active than traditionally prepared human Aβ oligomers, which contain SDS-stable trimers and tetramers, but are devoid of dimers. When incubated under tyrosine oxidizing conditions, synthetic human but not rodent Aβ1-42, the latter lacking tyrosine, acquires a marked increase (620 fold for EC50) in rod-inducing activity. Gel filtration of this preparation yielded two fractions containing SDS-stable dimers, trimers and tetramers. One, eluting at a similar volume to 7PA2 Aβd/t, had maximum activity at ~5 nM, whereas the other, eluting at the void volume (high-n state), lacked rod inducing activity at the same concentration. Fractions from 7PA2 medium containing Aβ monomers are not active, suggesting oxidized SDS-stable Aβ1-42 dimers in a low-n state are the most active rod-inducing species. Aβd/t-induced rods are predominantly localized to the dentate gyrus and mossy fiber tract, reach significance over controls within 2 h of treatment, and are reversible, disappearing by 24 h after Aβd/t washout. Overexpression of cofilin phosphatases increase rod formation when expressed alone and exacerbate rod formation when coupled with Aβd/t, whereas overexpression of a cofilin kinase inhibits Aβd/t-induced rod formation.
Conclusions: Together these data support a mechanism by which Aβd/t alters the actin cytoskeleton via effects on cofilin in neurons critical to learning and memory.
Figures
References
-
- Sisodia SS, Price DL. Role of the beta-amyloid protein in Alzheimer's disease. FASEB J. 1995;9:366–370. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
