

RIM determines Ca²+ channel density and vesicle docking at the presynaptic active zone
- PMID: 21262468
- PMCID: PMC3259453
- DOI: 10.1016/j.neuron.2010.12.014
RIM determines Ca²+ channel density and vesicle docking at the presynaptic active zone
Abstract
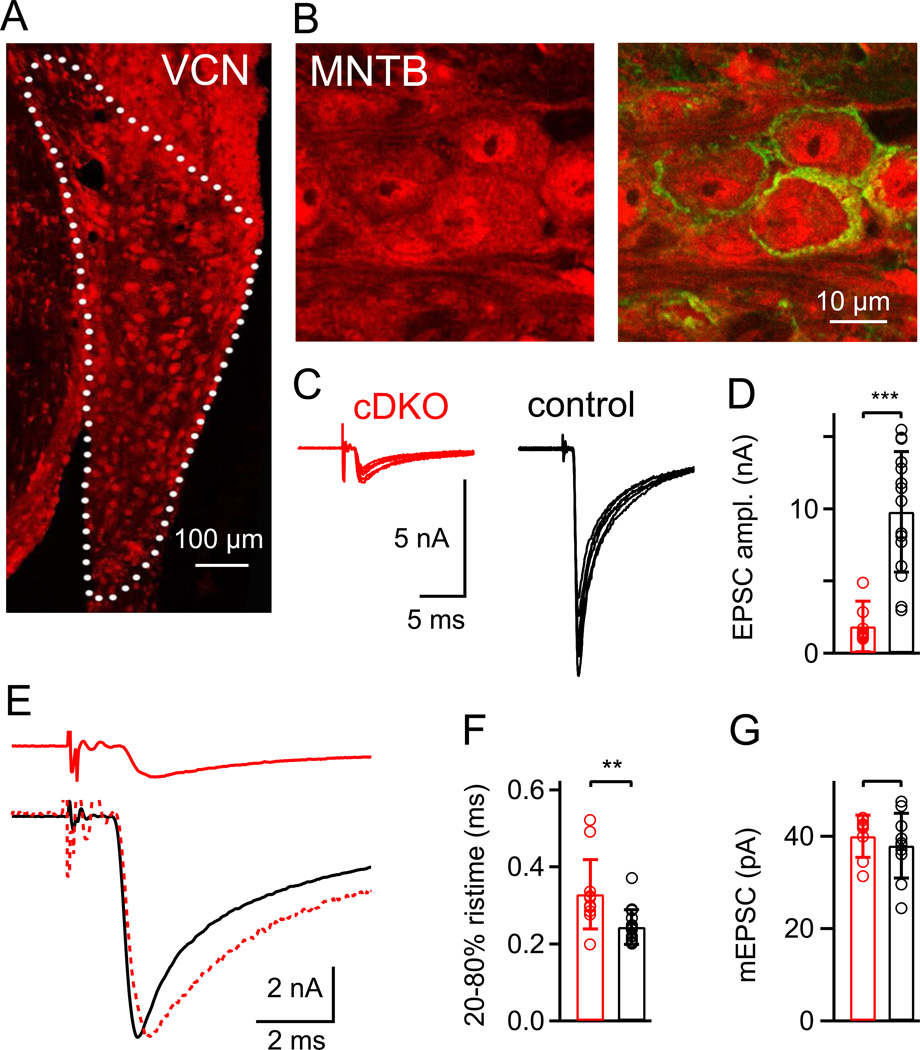
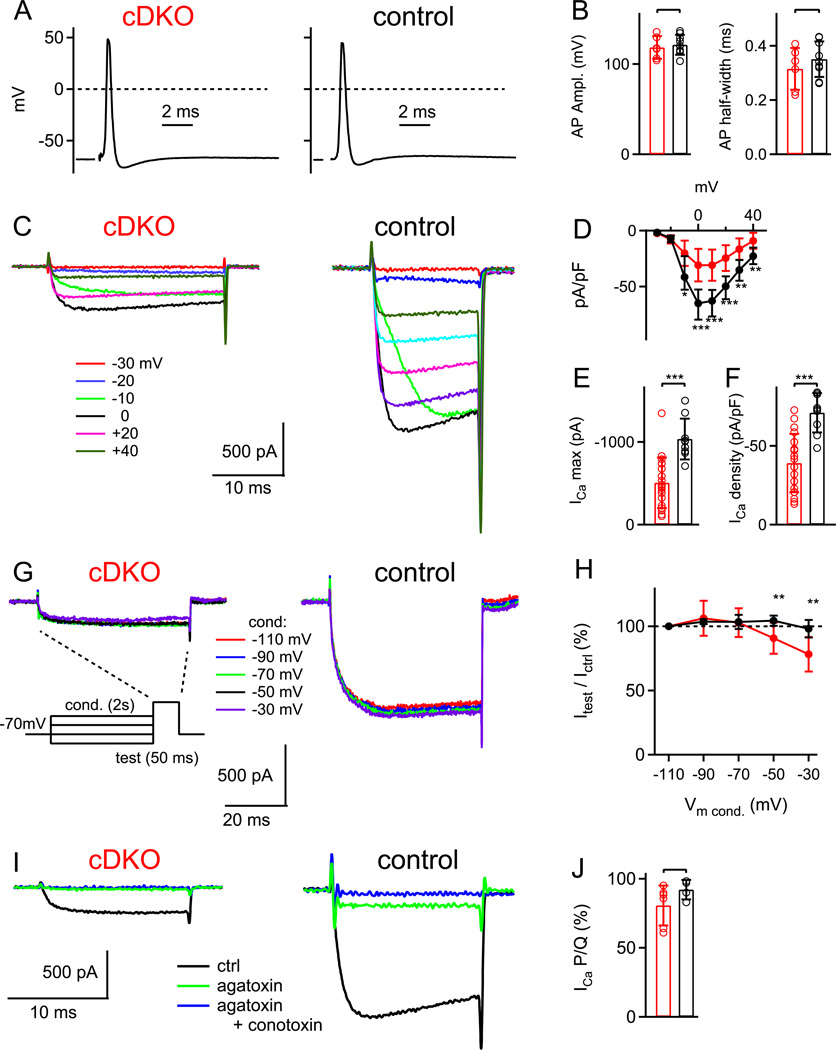
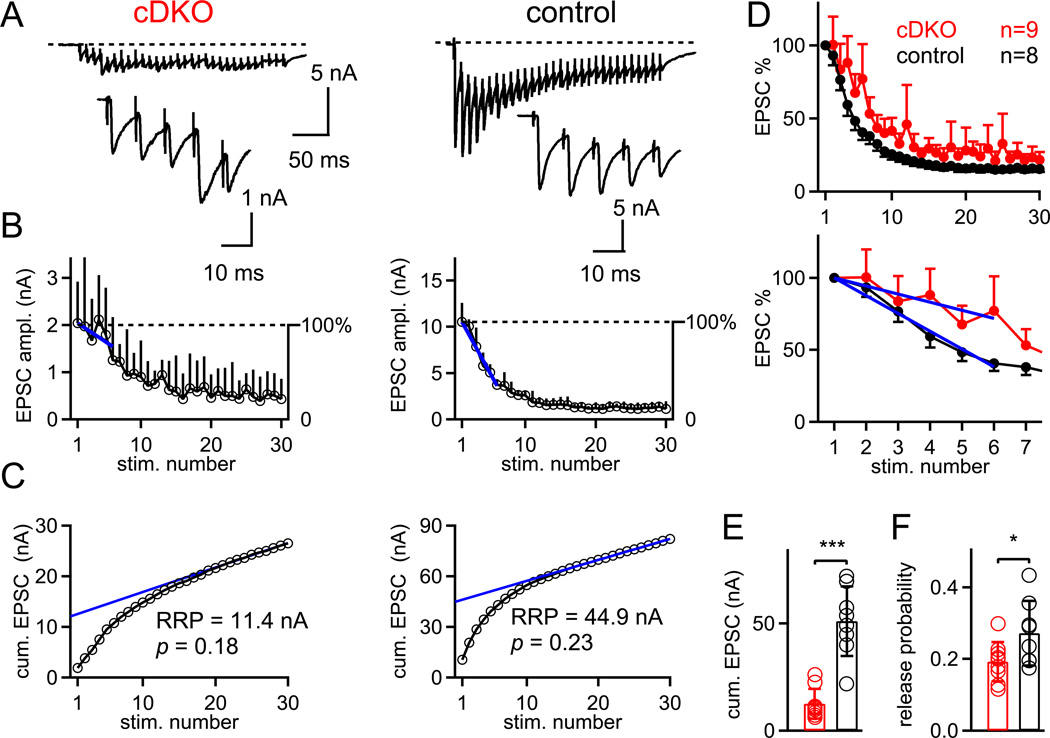
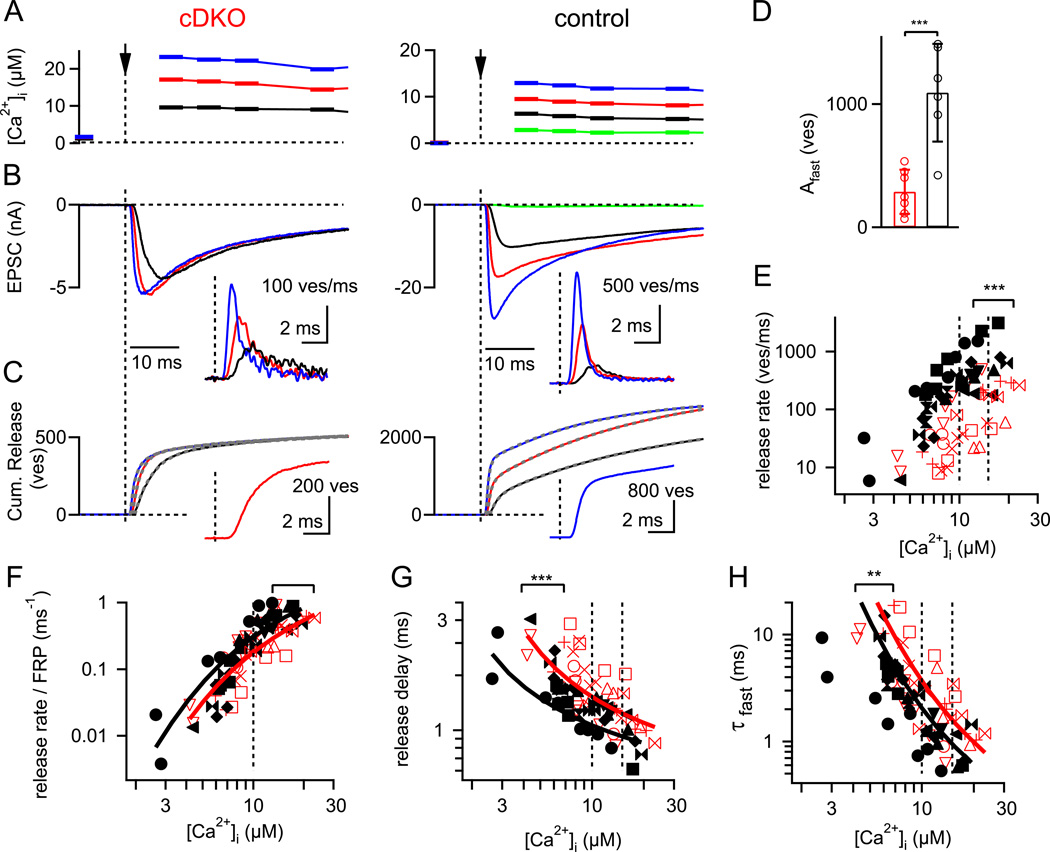
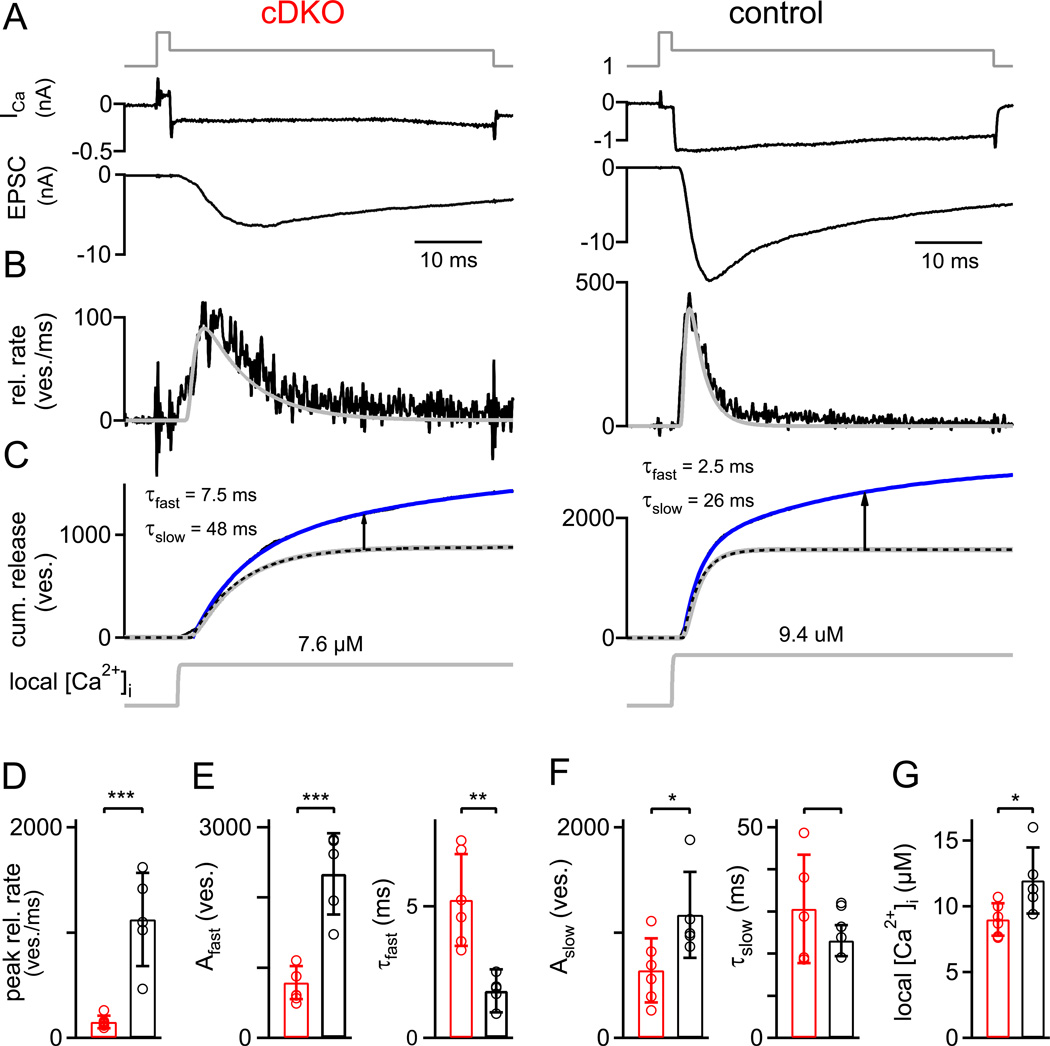
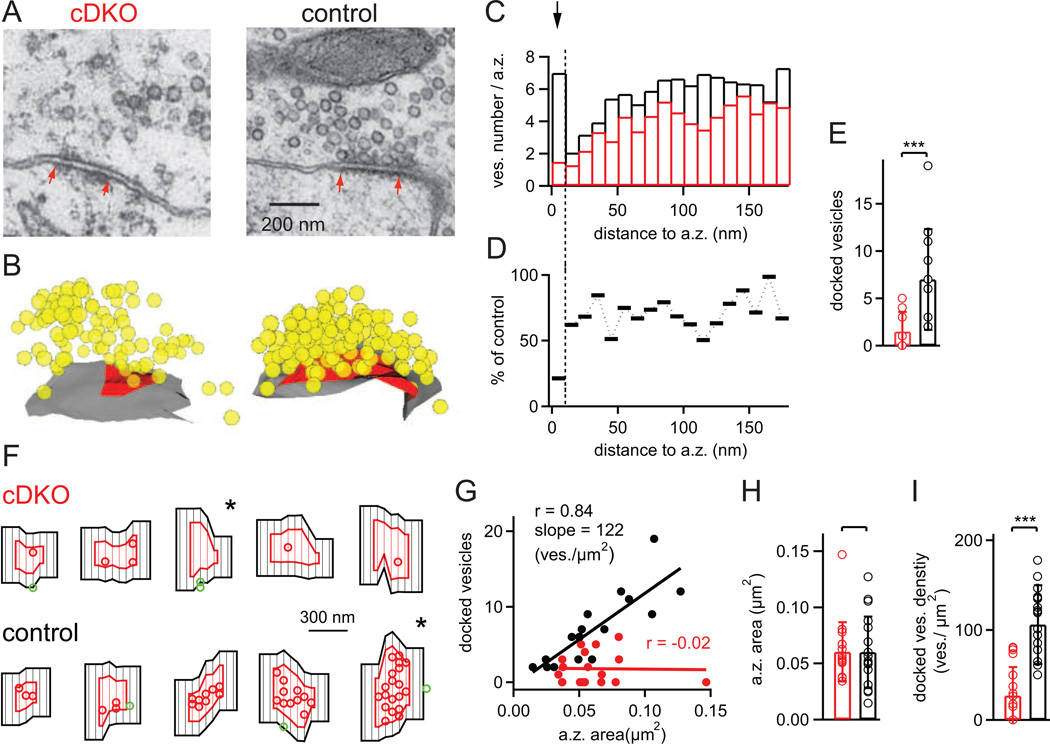
At presynaptic active zones, neurotransmitter release is initiated by the opening of voltage-gated Ca²+ channels close to docked vesicles. The mechanisms that enrich Ca²+ channels at active zones are, however, largely unknown, possibly because of the limited presynaptic accessibility of most synapses. Here, we have established a Cre-lox based conditional knockout approach at a presynaptically accessible central nervous system synapse, the calyx of Held, to directly study the functions of RIM proteins. Removal of all RIM1/2 isoforms strongly reduced the presynaptic Ca²+ channel density, revealing a role of RIM proteins in Ca²+ channel targeting. Removal of RIMs also reduced the readily releasable pool, paralleled by a similar reduction of the number of docked vesicles, and the Ca²+ channel-vesicle coupling was decreased. Thus, RIM proteins co-ordinately regulate key functions for fast transmitter release, enabling a high presynaptic Ca²+ channel density and vesicle docking at the active zone.
© 2011 Elsevier Inc. All rights reserved.
Figures
Comment in
-
The multiple faces of RIM.Neuron. 2011 Jan 27;69(2):185-7. doi: 10.1016/j.neuron.2011.01.010. Neuron. 2011. PMID: 21262457
References
-
- Augustin I, Rosenmund C, Südhof TC, Brose N. Munc13-1 is essential for fusion competence of glutamatergic synaptic vesicles. Nature. 1999;400:457–461. - PubMed
-
- Betz A, Thakur P, Junge HJ, Ashery U, Rhee JS, Scheuss V, Rosenmund C, Rettig J, Brose N. Functional interaction of the active zone proteins Munc13-1 and RIM1 in synaptic vesicle priming. Neuron. 2001;30:183–196. - PubMed
-
- Borst JG, Sakmann B. Calcium influx and transmitter release in a fast CNS synapse. Nature. 1996;383:431–434. - PubMed
-
- Bucurenciu I, Kulik A, Schwaller B, Frotscher M, Jonas P. Nanodomain coupling between Ca2+ channels and Ca2+ sensors promotes fast and efficient transmitter release at a cortical GABAergic synapse. Neuron. 2008;57:536–545. - PubMed
-
- Calakos N, Schoch S, Südhof TC, Malenka RC. Multiple roles for the active zone protein RIM1α in late stages of neurotransmitter release. Neuron. 2004;42:889–896. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
