

Tctex-1, a novel interaction partner of Rab3D, is required for osteoclastic bone resorption
- PMID: 21262767
- PMCID: PMC3135285
- DOI: 10.1128/MCB.00834-10
Tctex-1, a novel interaction partner of Rab3D, is required for osteoclastic bone resorption
Abstract
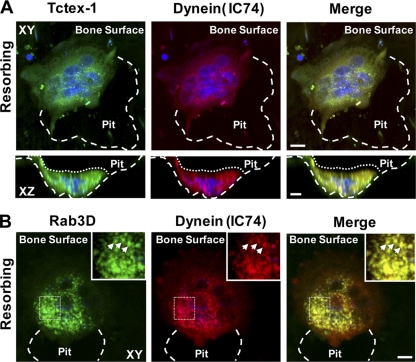
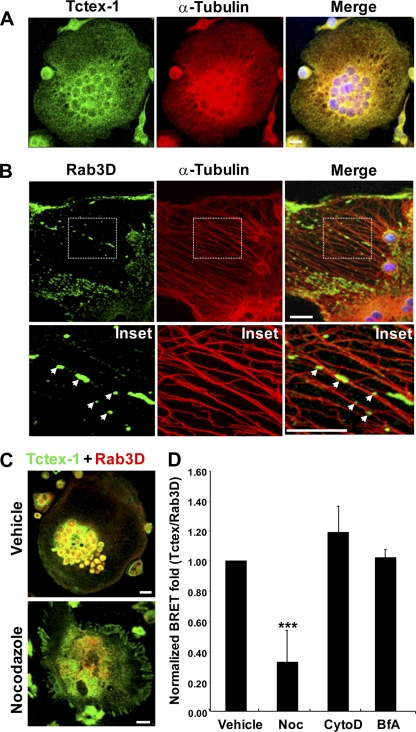
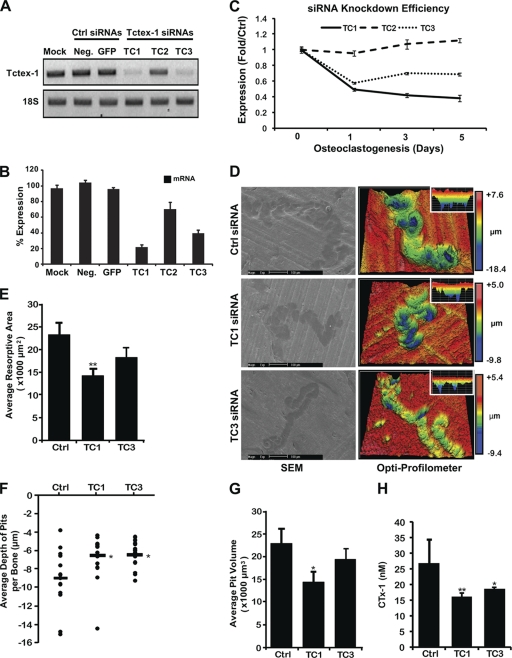
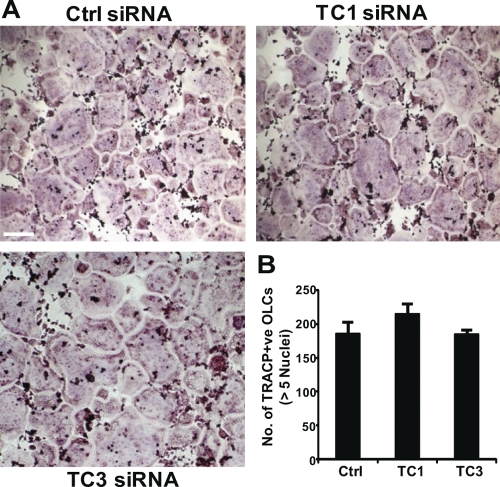
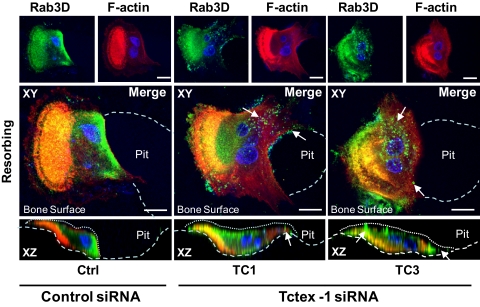
Vesicular transport along microtubules must be strictly regulated to sustain the unique structural and functional polarization of bone-resorbing osteoclasts. However, the molecular mechanisms bridging these vesicle-microtubule interactions remain largely obscure. Rab3D, a member of the Rab3 subfamily (Rab3A/B/C/D) of small exocytotic GTPases, represents a core component of the osteoclastic vesicle transport machinery. Here, we identify a new Rab3D-interacting partner, Tctex-1, a light chain of the cytoplasmic dynein microtubule motor complex, by a yeast two-hybrid screen. We demonstrate that Tctex-1 binds specifically to Rab3D in a GTP-dependent manner and co-occupies Rab3D-bearing vesicles in bone-resorbing osteoclasts. Furthermore, we provide evidence that Tctex-1 and Rab3D intimately associate with the dynein motor complex and microtubules in osteoclasts. Finally, targeted disruption of Tctex-1 by RNA interference significantly impairs bone resorption capacity and mislocalizes Rab3D vesicles in osteoclasts, attesting to the notion that components of the Rab3D-trafficking pathway contribute to the maintenance of osteoclastic resorptive function.
Figures
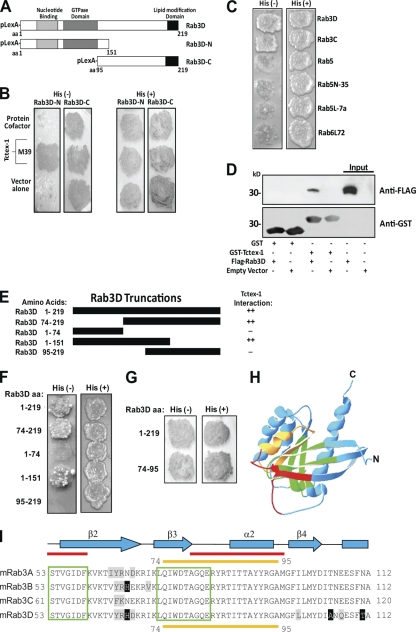
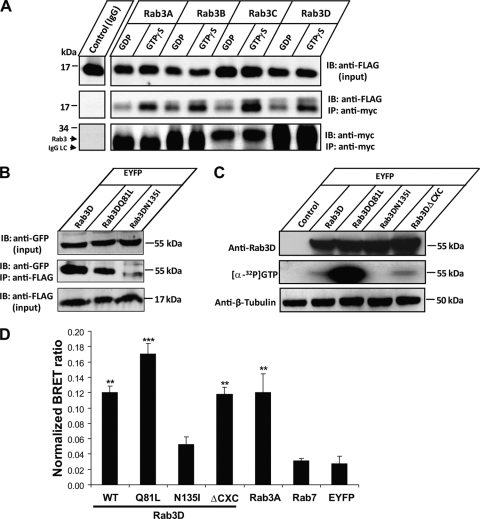
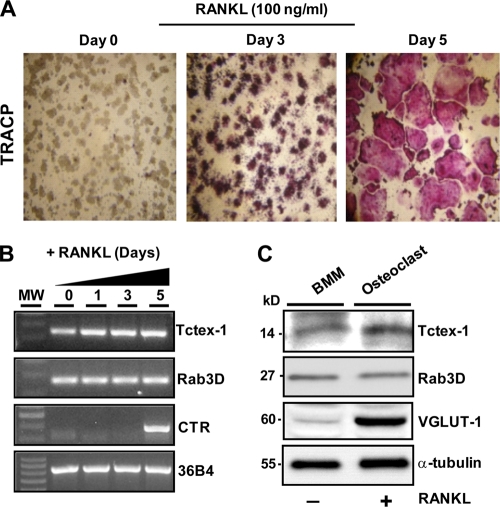
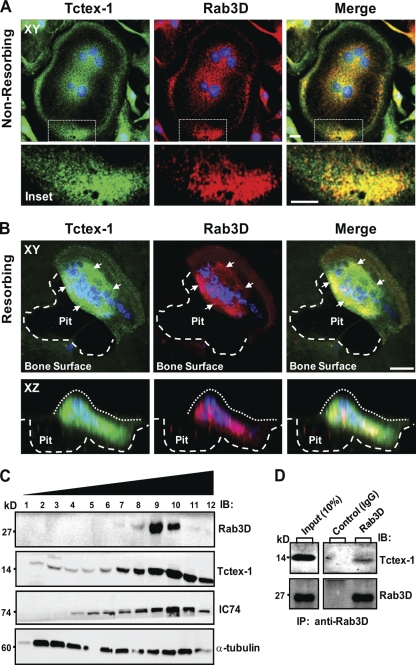
References
-
- Abu-Amer Y., Teitelbaum S. L., Chappel J. C., Schlesinger P., Ross F. P. 1999. Expression and regulation of RAB3 proteins in osteoclasts and their precursors. J. Bone Miner. Res. 14:1855–1860 - PubMed
-
- Campbell K. S., Cooper S., Dessing M., Yates S., Buder A. 1998. Interaction of p59fyn kinase with the dynein light chain, Tctex-1, and colocalization during cytokinesis. J. Immunol. 161:1728–1737 - PubMed
-
- Coxon F. P., Taylor A. 2008. Vesicular trafficking in osteoclasts. Semin. Cell Dev. Biol. 19:424–433 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases