

Human NDR kinases control G(1)/S cell cycle transition by directly regulating p21 stability
- PMID: 21262772
- PMCID: PMC3135299
- DOI: 10.1128/MCB.01216-10
Human NDR kinases control G(1)/S cell cycle transition by directly regulating p21 stability
Abstract
The G(1) phase of the cell cycle is an important integrator of internal and external cues, allowing a cell to decide whether to proliferate, differentiate, or die. Multiple protein kinases, among them the cyclin-dependent kinases (Cdks), control G(1)-phase progression and S-phase entry. With the regulation of apoptosis, centrosome duplication, and mitotic chromosome alignment downstream of the HIPPO pathway components MST1 and MST2, mammalian NDR kinases have been implicated to function in cell cycle-dependent processes. Although they are well characterized in terms of biochemical regulation and upstream signaling pathways, signaling mechanisms downstream of mammalian NDR kinases remain largely unknown. We identify here a role for human NDR in regulating the G(1)/S transition. In G(1) phase, NDR kinases are activated by a third MST kinase (MST3). Significantly, interfering with NDR and MST3 kinase expression results in G(1) arrest and subsequent proliferation defects. Furthermore, we describe the first downstream signaling mechanisms by which NDR kinases regulate cell cycle progression. Our findings suggest that NDR kinases control protein stability of the cyclin-Cdk inhibitor protein p21 by direct phosphorylation. These findings establish a novel MST3-NDR-p21 axis as an important regulator of G(1)/S progression of mammalian cells.
Figures
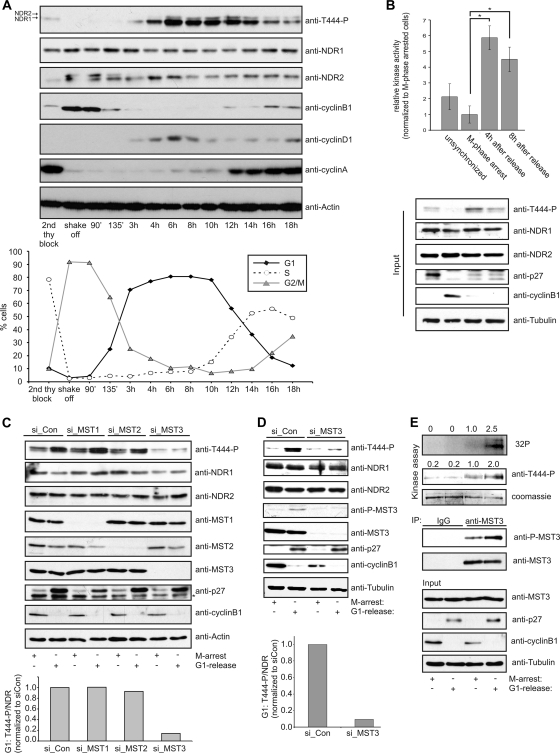
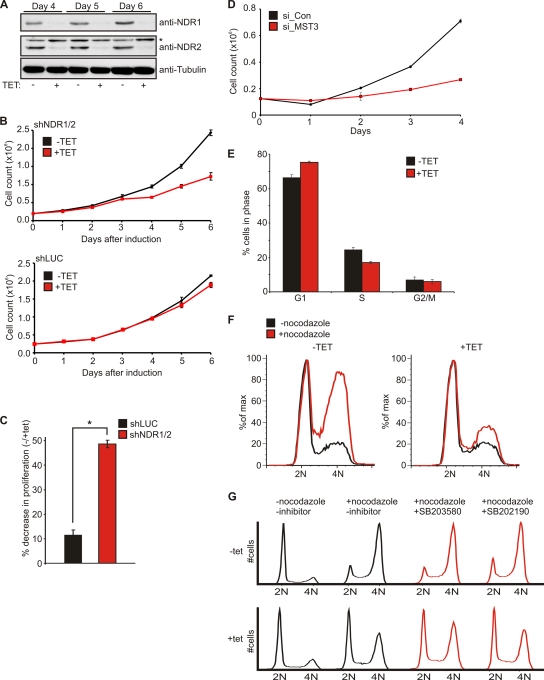
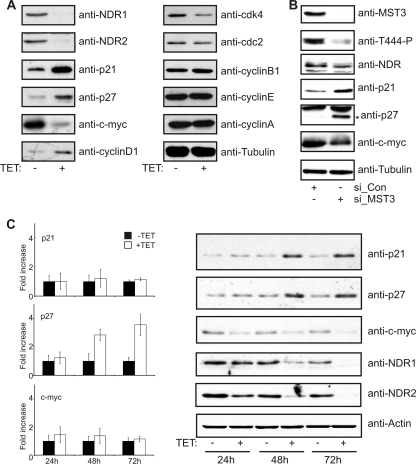
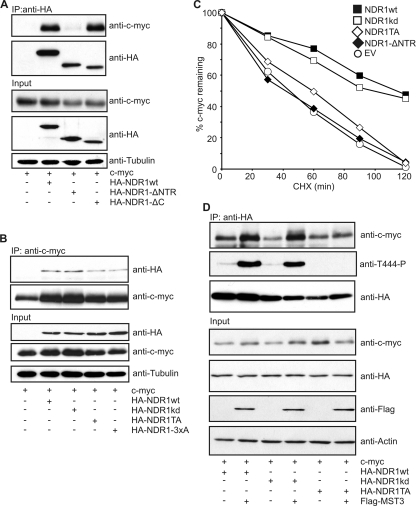
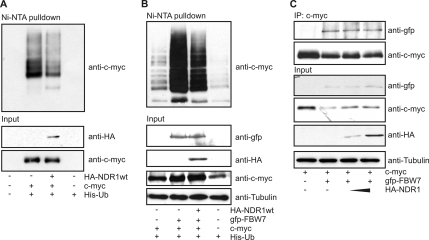
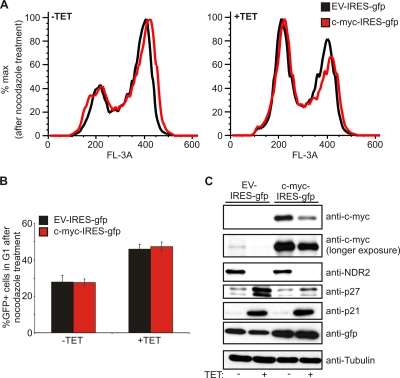
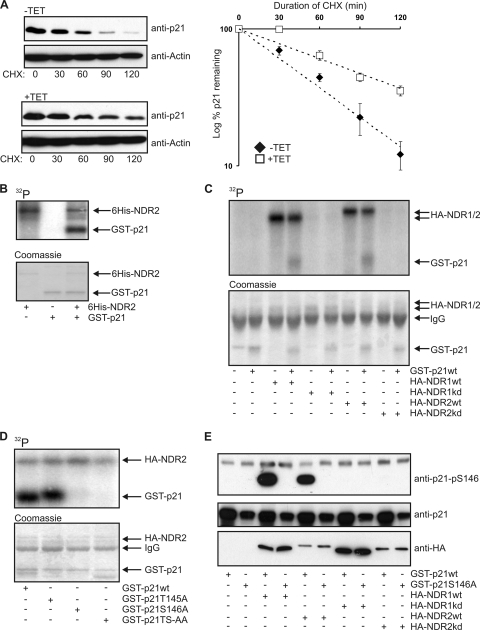
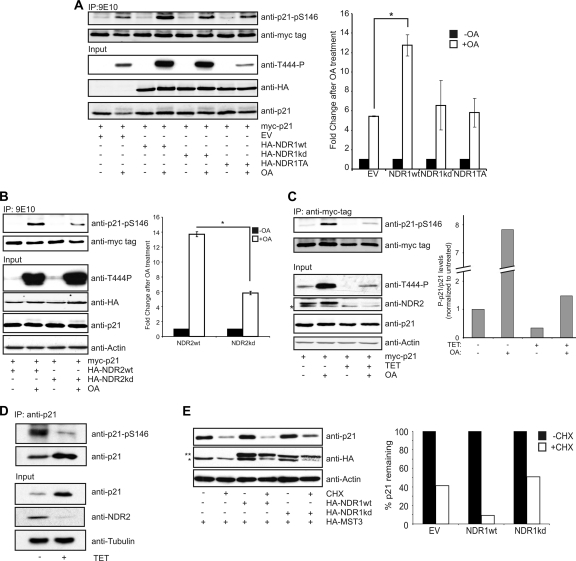
References
-
- Bichsel S. J., Tamaskovic R., Stegert M. R., Hemmings B. A. 2004. Mechanism of activation of NDR (nuclear Dbf2-related) protein kinase by the hMOB1 protein. J. Biol. Chem. 279:35228–35235 - PubMed
-
- Bothos J., Tuttle R. L., Ottey M., Luca F. C., Halazonetis T. D. 2005. Human LATS1 is a mitotic exit network kinase. Cancer Res. 65:6568–6575 - PubMed
-
- Chen C. B., Ng J. K., Choo P. H., Wu W., Porter A. G. 2009. Mammalian sterile 20-like kinase 3 (MST3) mediates oxidative-stress-induced cell death by modulating JNK activation. Biosci. Rep. 29:405–415 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous