

Casein kinase-2 mediates cell survival through phosphorylation and degradation of inositol hexakisphosphate kinase-2
- PMID: 21262846
- PMCID: PMC3038742
- DOI: 10.1073/pnas.1019381108
Casein kinase-2 mediates cell survival through phosphorylation and degradation of inositol hexakisphosphate kinase-2
Abstract
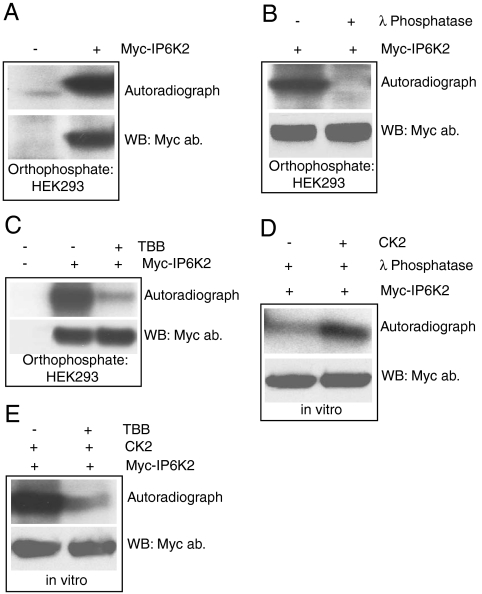
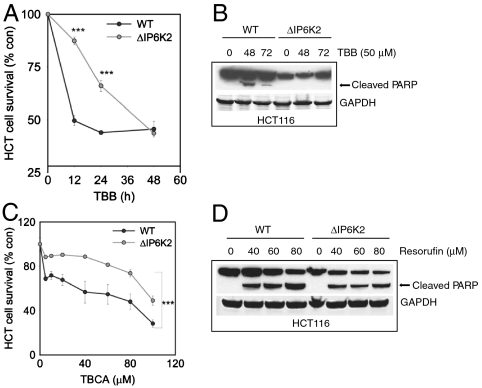
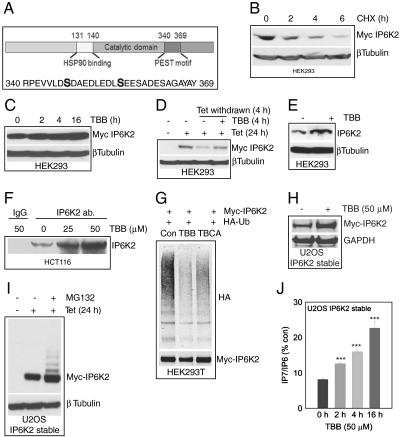
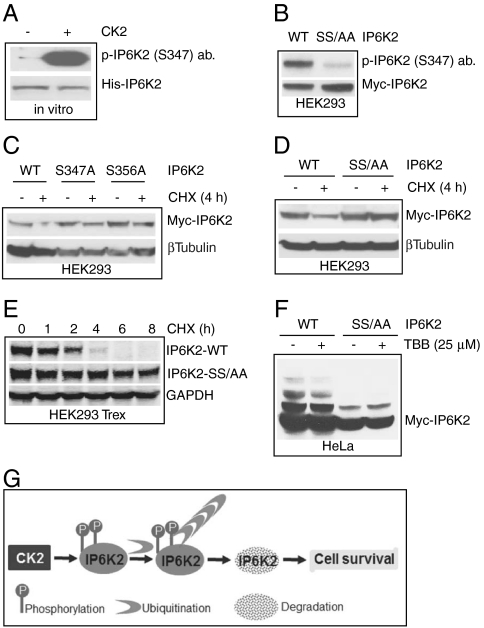
The inositol pyrophosphate, diphosphoinositol pentakisphosphate, regulates p53 and protein kinase Akt signaling, and its aberrant increase in cells has been implicated in apoptosis and insulin resistance. Inositol hexakisphosphate kinase-2 (IP6K2), one of the major inositol pyrophosphate synthesizing enzymes, mediates p53-linked apoptotic cell death. Casein kinase-2 (CK2) promotes cell survival and is upregulated in tumors. We show that CK2 mediated cell survival involves IP6K2 destabilization. CK2 physiologically phosphorylates IP6K2 at amino acid residues S347 and S356 contained within a PEST sequence, a consensus site for ubiquitination. HCT116 cells depleted of IP6K2 are resistant to cell death elicited by CK2 inhibitors. CK2 phosphorylation at the degradation motif of IP6K2 enhances its ubiquitination and subsequent degradation. IP6K2 mutants at the CK2 sites that are resistant to CK2 phosphorylation are metabolically stable.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Berridge MJ, Lipp P, Bootman MD. The versatility and universality of calcium signalling. Nat Rev Mol Cell Biol. 2000;1:11–21. - PubMed
-
- Hand CE, Honek JF. Phosphate transfer from inositol pyrophosphates InsP5PP and InsP4(PP)2: a semi-empirical investigation. Bioorg Med Chem Lett. 2007;17:183–188. - PubMed
-
- Mulugu S, et al. A conserved family of enzymes that phosphorylate inositol hexakisphosphate. Science. 2007;316:106–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
