

The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host
- PMID: 21262960
- PMCID: PMC3039851
- DOI: 10.1084/jem.20101488
The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host
Abstract
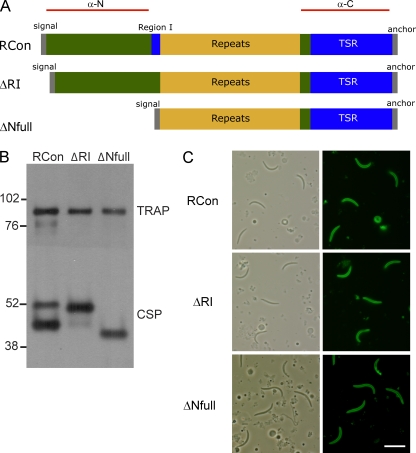
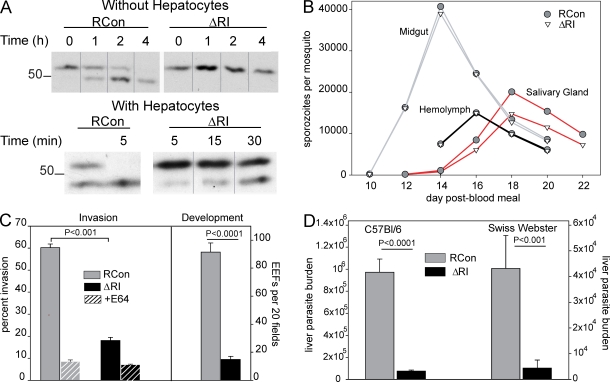
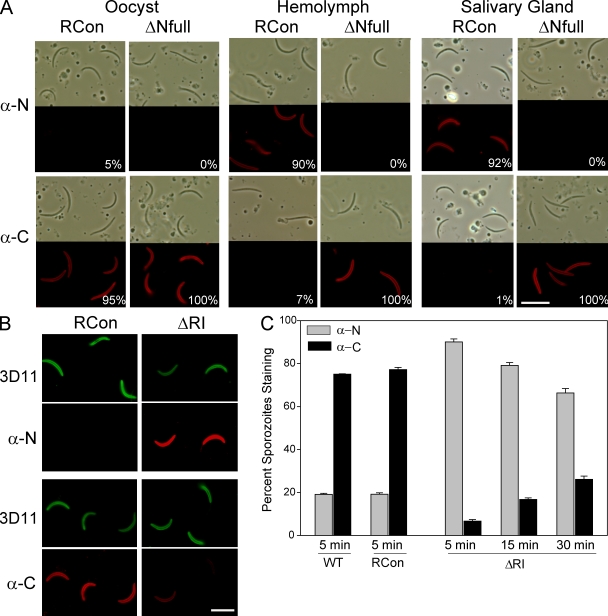
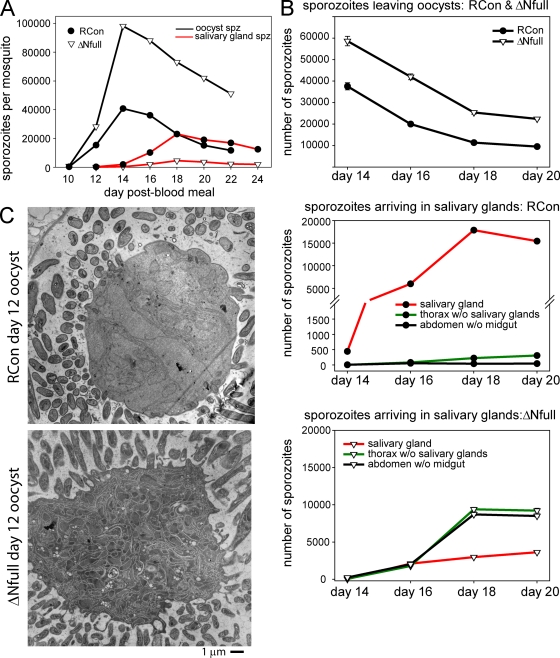
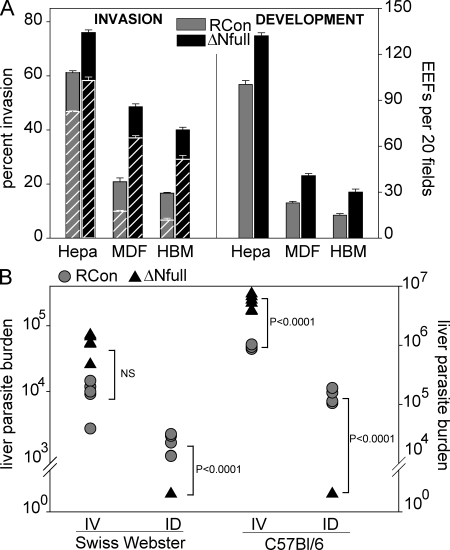
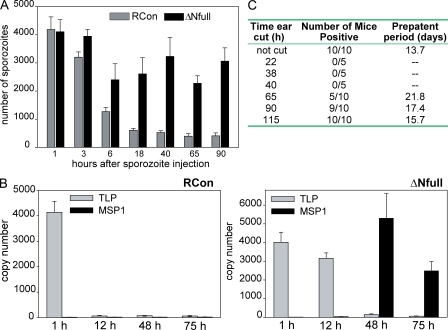
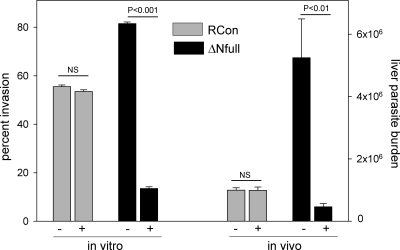
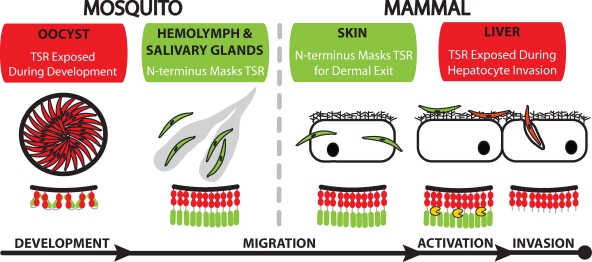
Plasmodium sporozoites make a remarkable journey from the mosquito midgut to the mammalian liver. The sporozoite's major surface protein, circumsporozoite protein (CSP), is a multifunctional protein required for sporozoite development and likely mediates several steps of this journey. In this study, we show that CSP has two conformational states, an adhesive conformation in which the C-terminal cell-adhesive domain is exposed and a nonadhesive conformation in which the N terminus masks this domain. We demonstrate that the cell-adhesive domain functions in sporozoite development and hepatocyte invasion. Between these two events, the sporozoite must travel from the mosquito midgut to the mammalian liver, and N-terminal masking of the cell-adhesive domain maintains the sporozoite in a migratory state. In the mammalian host, proteolytic cleavage of CSP regulates the switch to an adhesive conformation, and the highly conserved region I plays a critical role in this process. If the CSP domain architecture is altered such that the cell-adhesive domain is constitutively exposed, the majority of sporozoites do not reach their target organs, and in the mammalian host, they initiate a blood stage infection directly from the inoculation site. These data provide structure-function information relevant to malaria vaccine development.
Figures
References
-
- Amino R., Giovannini D., Thiberge S., Gueirard P., Boisson B., Dubremetz J.F., Prévost M.C., Ishino T., Yuda M., Ménard R. 2008. Host cell traversal is important for progression of the malaria parasite through the dermis to the liver. Cell Host Microbe. 3:88–96 10.1016/j.chom.2007.12.007 - DOI - PubMed
