

Protein-tyrosine phosphatase DEP-1 controls receptor tyrosine kinase FLT3 signaling
- PMID: 21262971
- PMCID: PMC3064147
- DOI: 10.1074/jbc.M110.205021
Protein-tyrosine phosphatase DEP-1 controls receptor tyrosine kinase FLT3 signaling
Abstract
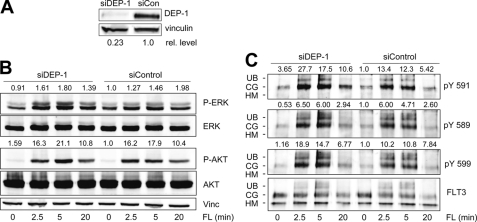
Fms-like tyrosine kinase 3 (FLT3) plays an important role in hematopoietic differentiation, and constitutively active FLT3 mutant proteins contribute to the development of acute myeloid leukemia. Little is known about the protein-tyrosine phosphatases (PTP) affecting the signaling activity of FLT3. To identify such PTP, myeloid cells expressing wild type FLT3 were infected with a panel of lentiviral pseudotypes carrying shRNA expression cassettes targeting different PTP. Out of 20 PTP tested, expressed in hematopoietic cells, or presumed to be involved in oncogenesis or tumor suppression, DEP-1 (PTPRJ) was identified as a PTP negatively regulating FLT3 phosphorylation and signaling. Stable 32D myeloid cell lines with strongly reduced DEP-1 levels showed site-selective hyperphosphorylation of FLT3. In particular, the sites pTyr-589, pTyr-591, and pTyr-842 involved in the FLT3 ligand (FL)-mediated activation of FLT3 were hyperphosphorylated the most. Similarly, acute depletion of DEP-1 in the human AML cell line THP-1 caused elevated FLT3 phosphorylation. Direct interaction of DEP-1 and FLT3 was demonstrated by "substrate trapping" experiments showing association of DEP-1 D1205A or C1239S mutant proteins with FLT3 by co-immunoprecipitation. Moreover, activated FLT3 could be dephosphorylated by recombinant DEP-1 in vitro. Enhanced FLT3 phosphorylation in DEP-1-depleted cells was accompanied by enhanced FLT3-dependent activation of ERK and cell proliferation. Stable overexpression of DEP-1 in 32D cells and transient overexpression with FLT3 in HEK293 cells resulted in reduction of FL-mediated FLT3 signaling activity. Furthermore, FL-stimulated colony formation of 32D cells expressing FLT3 in methylcellulose was induced in response to shRNA-mediated DEP-1 knockdown. This transforming effect of DEP-1 knockdown was consistent with a moderately increased activation of STAT5 upon FL stimulation but did not translate into myeloproliferative disease formation in the 32D-C3H/HeJ mouse model. The data indicate that DEP-1 is negatively regulating FLT3 signaling activity and that its loss may contribute to but is not sufficient for leukemogenic cell transformation.
Figures





References
-
- Stirewalt D. L., Radich J. P. (2003) Nat. Rev. Cancer 3, 650–665 - PubMed
-
- Schmidt-Arras D., Schwäble J., Böhmer F. D., Serve H. (2004) Curr. Pharm. Des. 10, 1867–1883 - PubMed
-
- Gilliland D. G., Griffin J. D. (2002) Blood 100, 1532–1542 - PubMed
-
- Thiede C., Steudel C., Mohr B., Schaich M., Schäkel U., Platzbecker U., Wermke M., Bornhäuser M., Ritter M., Neubauer A., Ehninger G., Illmer T. (2002) Blood 99, 4326–4335 - PubMed
-
- Small D. (2006) Hematology Am. Soc. Hematol. Educ. Program, 178–184 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
