

Phosphorylation regulates c-Myc's oncogenic activity in the mammary gland
- PMID: 21266350
- PMCID: PMC3077809
- DOI: 10.1158/0008-5472.CAN-10-1032
Phosphorylation regulates c-Myc's oncogenic activity in the mammary gland
Abstract
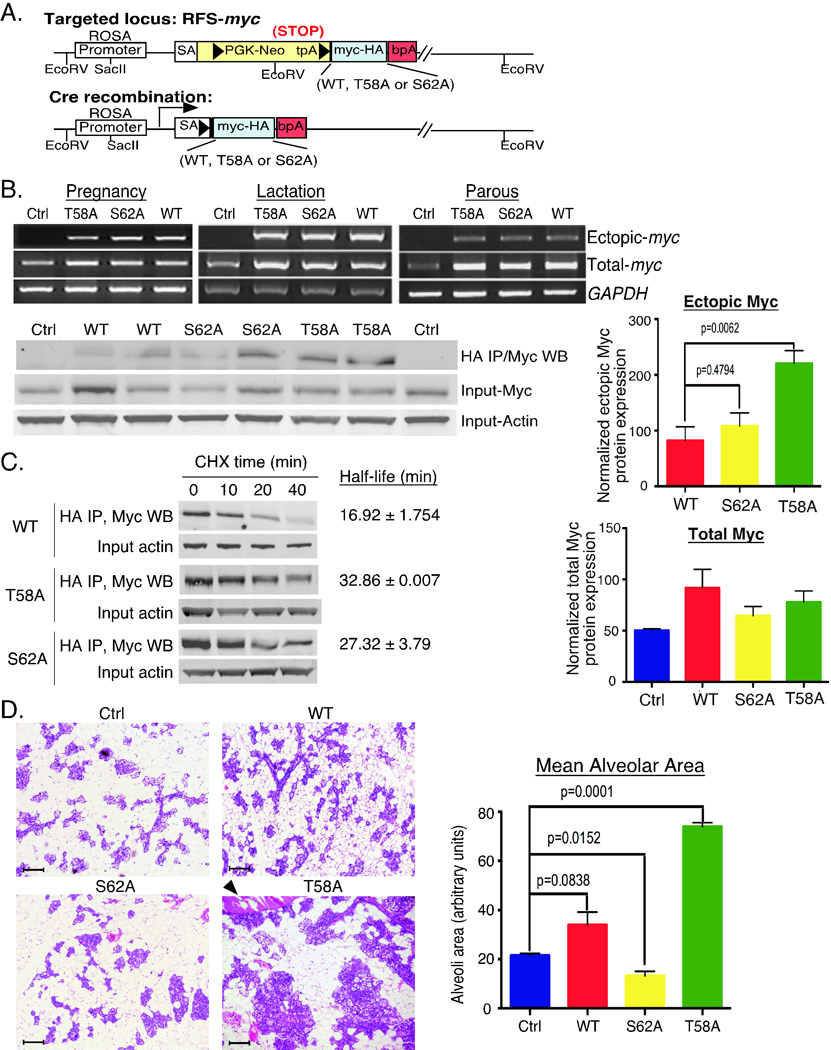
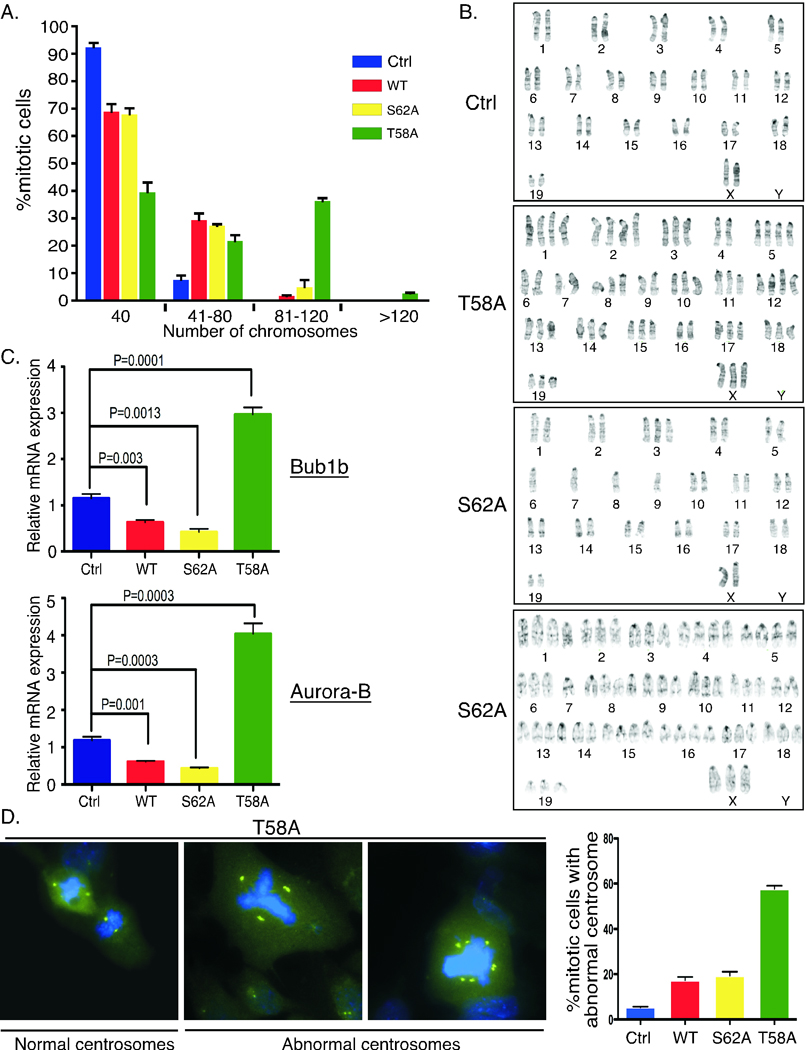
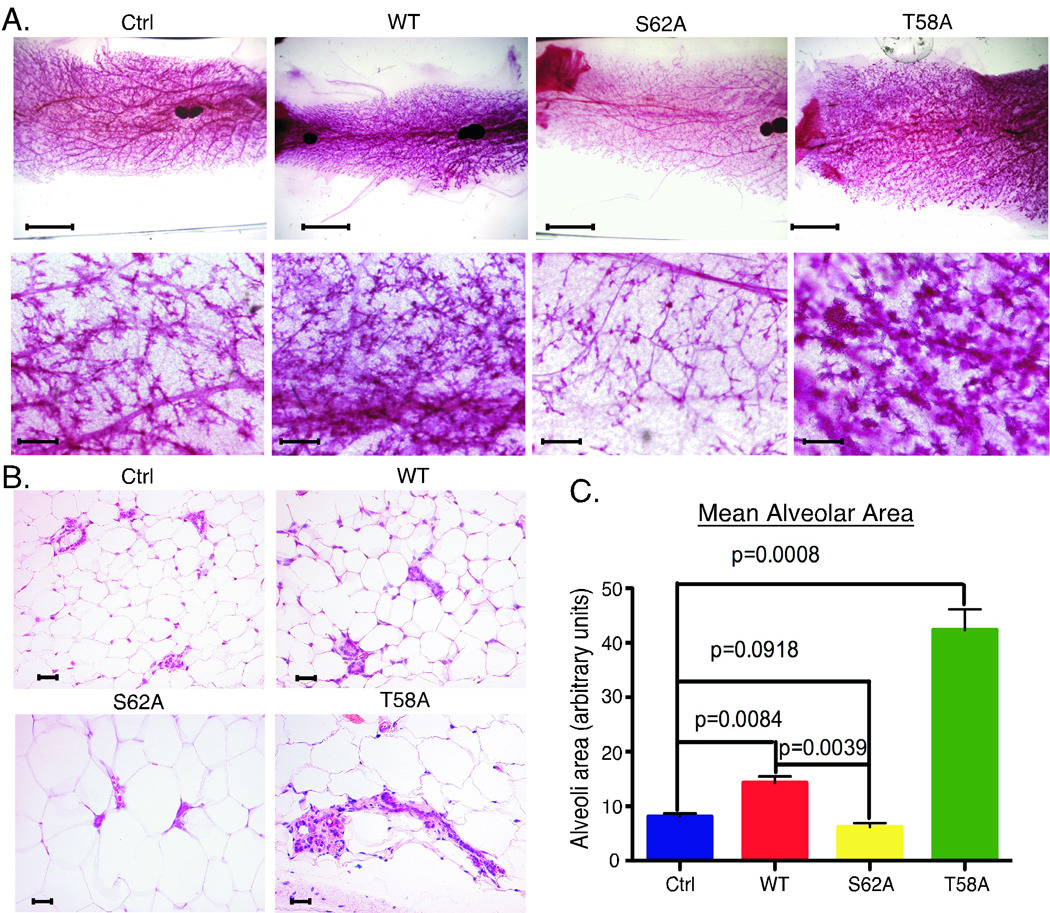
Expression of the c-Myc oncoprotein is affected by conserved threonine 58 (T58) and serine 62 (S62) phosphorylation sites that help to regulate c-Myc protein stability, and altered ratios of T58 and S62 phosphorylation have been observed in human cancer. Here, we report the development of 3 unique c-myc knock-in mice that conditionally express either c-Myc(WT) or the c-Myc(T58A) or c-Myc(S62A) phosphorylation mutant from the constitutively active ROSA26 locus in response to Cre recombinase to study the role of these phosphorylation sites in vivo. Using a mammary-specific Cre model, we found that expression of c-Myc(WT) resulted in increased mammary gland density, but normal morphology and no tumors at the level expressed from the ROSA promoter. In contrast, c-Myc(T58A) expression yielded enhanced mammary gland density, hyperplastic foci, cellular dysplasia, and mammary carcinoma, associated with increased genomic instability and suppressed apoptosis relative to c-Myc(WT). Alternatively, c-Myc(S62A) expression reduced mammary gland density relative to control glands, and this was associated with increased genomic instability and normal apoptotic function. Our results indicate that specific activities of c-Myc are differentially affected by T58 and S62 phosphorylation. This model provides a robust platform to interrogate the role that these phosphorylation sites play in c-Myc function during development and tumorigenesis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Meyer N, Penn LZ. Reflecting on 25 years with MYC. Nat Rev Cancer. 2008;8:976–990. - PubMed
-
- Nesbit CE, Tersak JM, Prochownik EV. MYC oncogenes and human neoplastic disease. Oncogene. 1999;18:3004–3016. - PubMed
-
- D'Cruz CM, Gunther EJ, Boxer RB, et al. c-MYC induces mammary by means of a preferred pathway involving spontaneous Kras2 mutations. Nat Med. 2001;7:235–239. - PubMed
-
- Pelengaris S, Khan M, Evan G. c-MYC: more than just a matter of life and death. Nat Rev Cancer. 2002;2:764–776. - PubMed
-
- Khan M, Evan G, Pelengaris S. Suppression of Myc-induced apoptosis in beta cells exposes multiple oncogenic properties of Myc and triggers carcinogenic progression. Nature Reviews Cancer. 2002;2:764–776. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
