

Punctuated actin contractions during convergent extension and their permissive regulation by the non-canonical Wnt-signaling pathway
- PMID: 21266466
- PMCID: PMC3031374
- DOI: 10.1242/jcs.067579
Punctuated actin contractions during convergent extension and their permissive regulation by the non-canonical Wnt-signaling pathway
Abstract
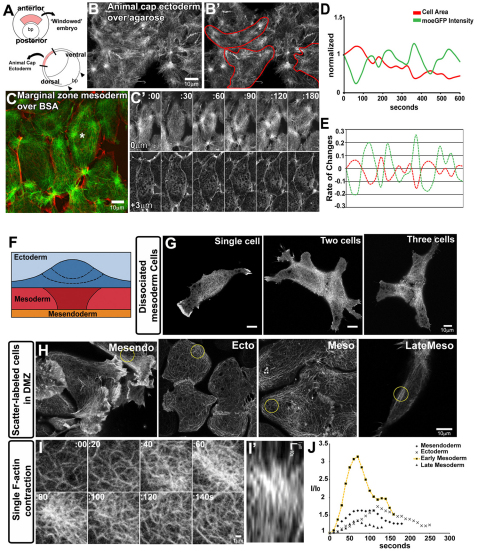
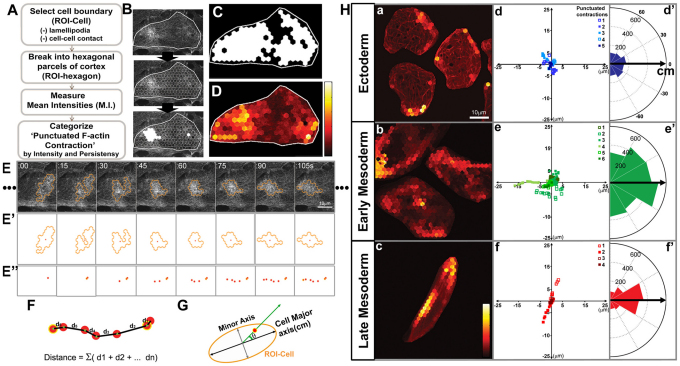
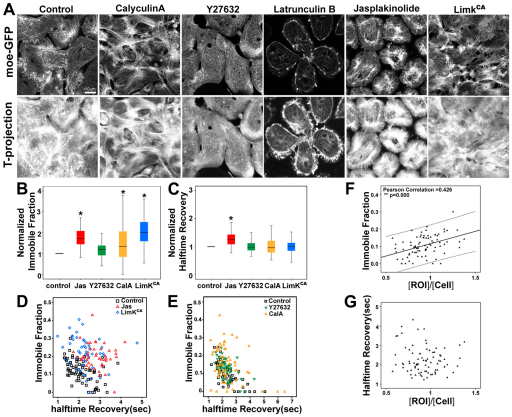
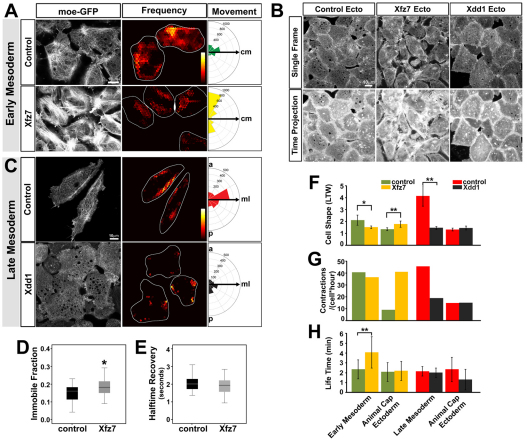
Actomyosin networks linked to the micro-environment through the plasma membrane are thought to be key players in regulating cell behaviors within multicellular tissues, such as converging and extending mesoderm. Here, we observe the dynamics of actin contractions called 'punctuated actin contractions' in the mid-cell body of embryonic mesenchymal cells in the mesoderm. These contraction dynamics are a common feature of Xenopus embryonic tissues and are important for cell shape changes during morphogenesis. Quantitative morphological analysis of these F-actin dynamics indicates that frequent and aligned movements of multiple actin contractions accompany mesoderm cells as they intercalate and elongate. Using inhibitors combined with fluorescence recovery after photobleaching (FRAP) analysis, we find that the dynamics of actin contractions are regulated by both myosin contractility and F-actin polymerization. Furthermore, we find that the non-canonical Wnt-signaling pathway permissively regulates levels of punctuated actin contractions. Overexpression of Xfz7 (Fzd7) can induce early maturation of actin contractions in mesoderm and produce mesoderm-like actin contractions in ectoderm cells. By contrast, expression of the dominant-negative Xenopus disheveled construct Xdd1 blocks the progression of actin contractions into their late mesoderm dynamics but has no effect in ectoderm. Our study reveals punctuated actin contractions within converging and extending mesoderm and uncovers a permissive role for non-canonical Wnt-signaling, myosin contractility and F-actin polymerization in regulating these dynamics.
Figures
References
-
- Abramoff M. D., Magelhaes P. J., Ram S. J. (2004). Image processing with ImageJ. Biophotonics International 11, 36-42
-
- Blalock H. M. (1972). Social Statistics. New York: McGraw-Hill;
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
