

SecM-stalled ribosomes adopt an altered geometry at the peptidyl transferase center
- PMID: 21267063
- PMCID: PMC3022528
- DOI: 10.1371/journal.pbio.1000581
SecM-stalled ribosomes adopt an altered geometry at the peptidyl transferase center
Abstract
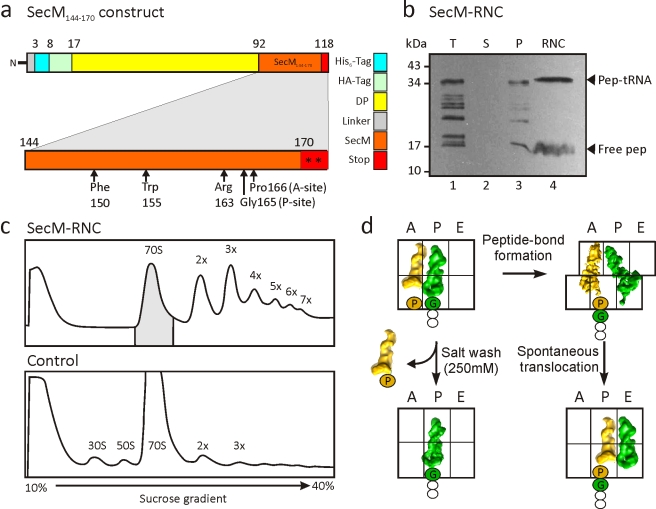
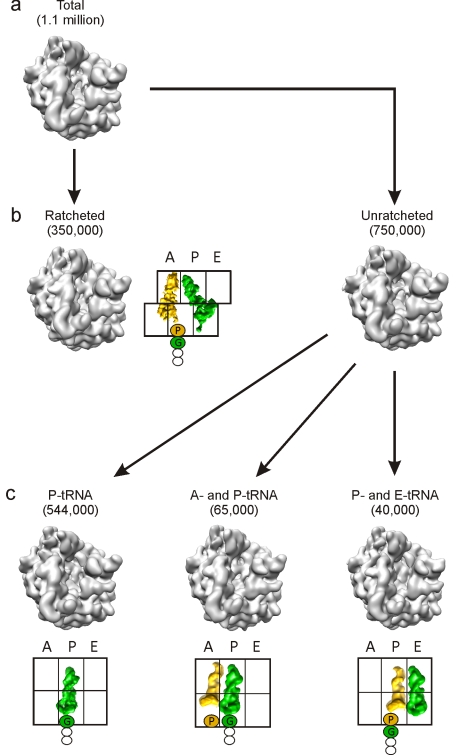
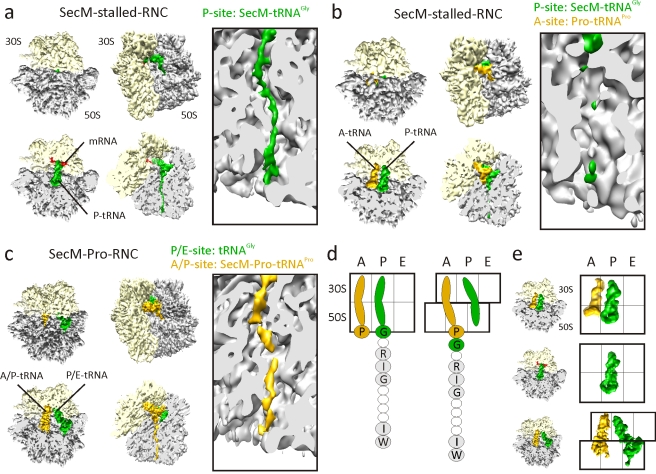
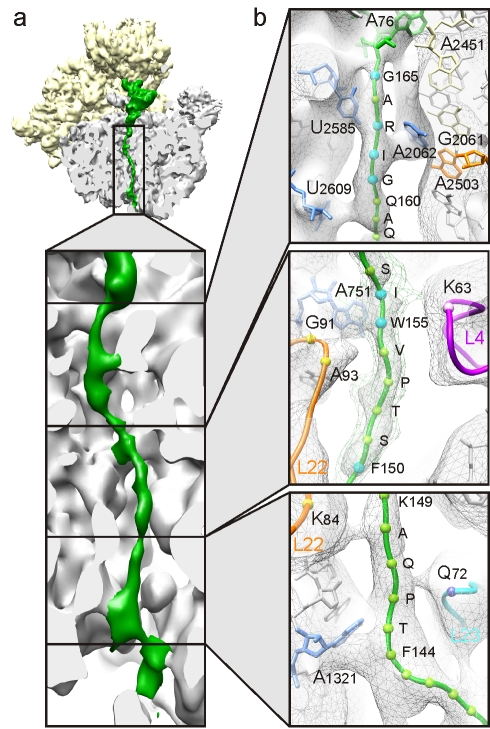
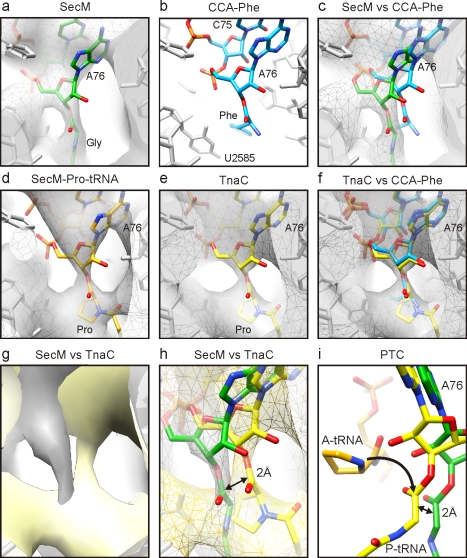
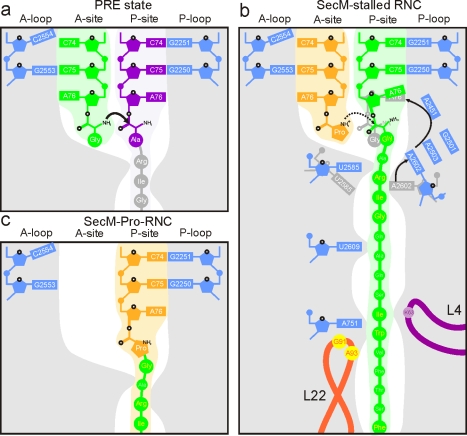
As nascent polypeptide chains are synthesized, they pass through a tunnel in the large ribosomal subunit. Interaction between specific nascent chains and the ribosomal tunnel is used to induce translational stalling for the regulation of gene expression. One well-characterized example is the Escherichia coli SecM (secretion monitor) gene product, which induces stalling to up-regulate translation initiation of the downstream secA gene, which is needed for protein export. Although many of the key components of SecM and the ribosomal tunnel have been identified, understanding of the mechanism by which the peptidyl transferase center of the ribosome is inactivated has been lacking. Here we present a cryo-electron microscopy reconstruction of a SecM-stalled ribosome nascent chain complex at 5.6 Å. While no cascade of rRNA conformational changes is evident, this structure reveals the direct interaction between critical residues of SecM and the ribosomal tunnel. Moreover, a shift in the position of the tRNA-nascent peptide linkage of the SecM-tRNA provides a rationale for peptidyl transferase center silencing, conditional on the simultaneous presence of a Pro-tRNA(Pro) in the ribosomal A-site. These results suggest a distinct allosteric mechanism of regulating translational elongation by the SecM stalling peptide.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Polacek N, Mankin A. S. The ribosomal peptidyl transferase center: structure, function, evolution, inhibition. Crit Rev Biochem Mol Biol. 2005;40:285–311. - PubMed
-
- Bhushan S, Gartmann M, Halic M, Armache J. P, Jarasch A, et al. Alpha-helical nascent polypeptide chains visualized within distinct regions of the ribosomal exit tunnel. Nat Struct Mol Biol. 2010;17:313–317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
