

Nonrenewal spike train statistics: causes and functional consequences on neural coding
- PMID: 21267548
- PMCID: PMC4529317
- DOI: 10.1007/s00221-011-2553-y
Nonrenewal spike train statistics: causes and functional consequences on neural coding
Erratum in
-
Erratum to: Nonrenewal spike train statistics: causes and functional consequences on neural coding.Exp Brain Res. 2011 May;210(3-4):373-5. doi: 10.1007/s00221-011-2639-6. Exp Brain Res. 2011. PMID: 21451983 Free PMC article. No abstract available.
Abstract
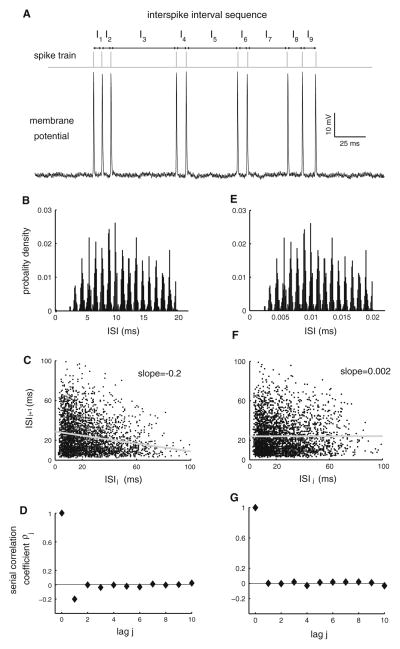
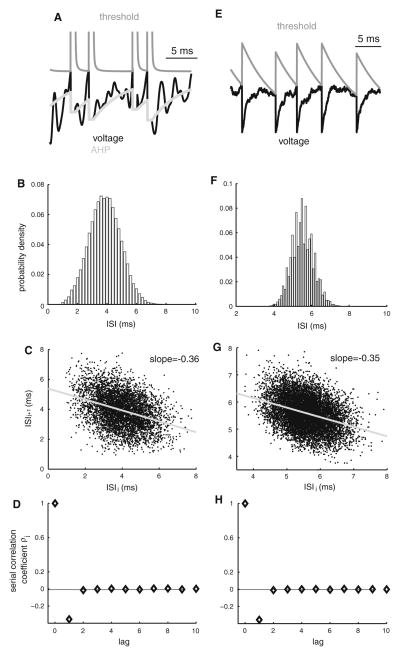
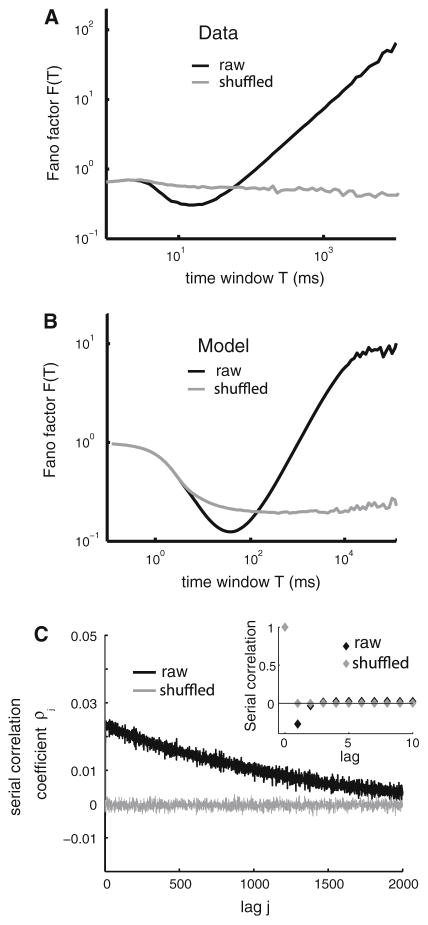
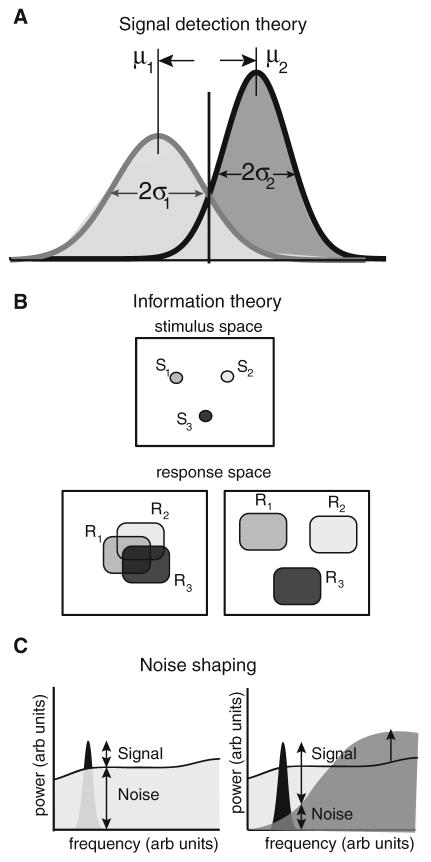
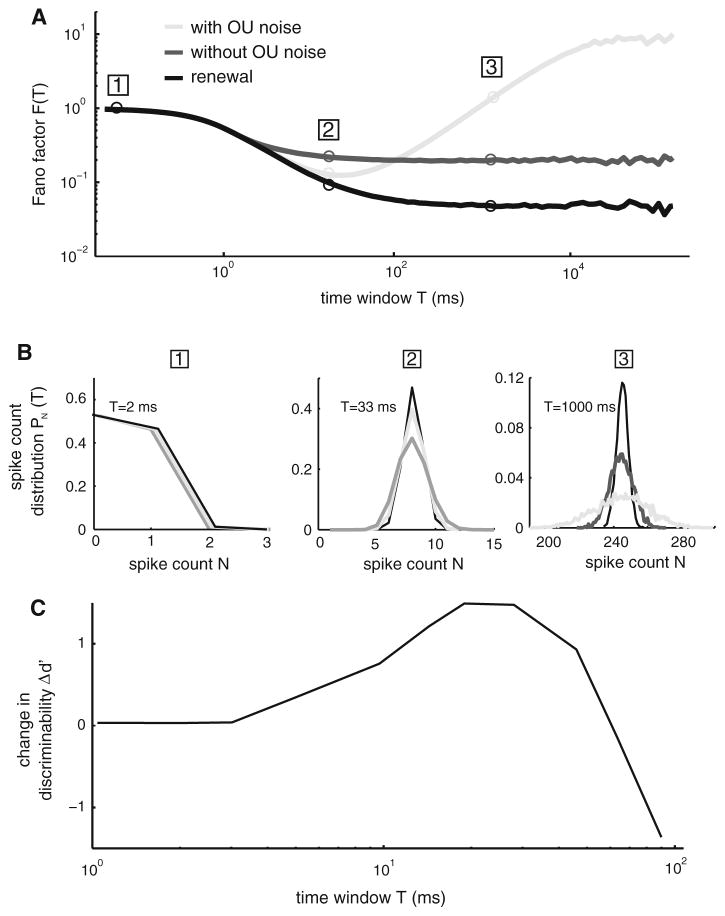
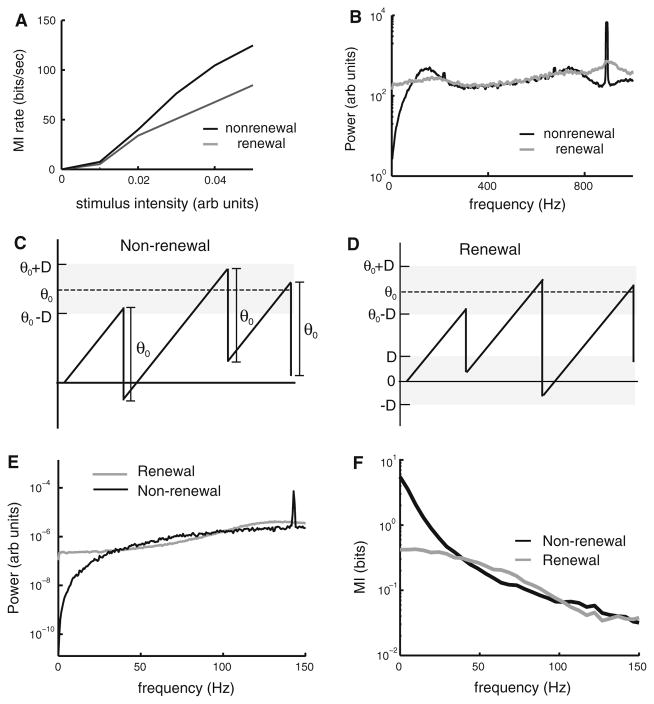
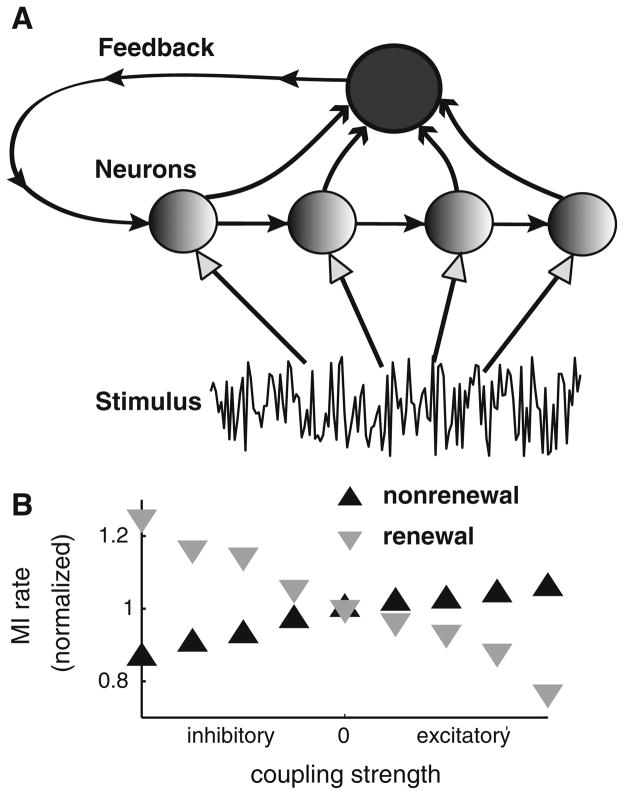
Many neurons display significant patterning in their spike trains (e.g. oscillations, bursting), and there is accumulating evidence that information is contained in these patterns. In many cases, this patterning is caused by intrinsic mechanisms rather than external signals. In this review, we focus on spiking activity that displays nonrenewal statistics (i.e. memory that persists from one firing to the next). Such statistics are seen in both peripheral and central neurons and appear to be ubiquitous in the CNS. We review the principal mechanisms that can give rise to nonrenewal spike train statistics. These are separated into intrinsic mechanisms such as relative refractoriness and network mechanisms such as coupling with delayed inhibitory feedback. Next, we focus on the functional roles for nonrenewal spike train statistics. These can either increase or decrease information transmission. We also focus on how such statistics can give rise to an optimal integration timescale at which spike train variability is minimal and how this might be exploited by sensory systems to maximize the detection of weak signals. We finish by pointing out some interesting future directions for research in this area. In particular, we explore the interesting possibility that synaptic dynamics might be matched with the nonrenewal spiking statistics of presynaptic spike trains in order to further improve information transmission.
Figures
References
-
- Abeles M. Corticonics. Cambridge University Press; Cambridge: 1991.
-
- Averbeck BB, Lee D. Effects of noise correlations on information encoding and decoding. J Neurophysiol. 2006;95:3633–3644. - PubMed
-
- Averbeck BB, Latham PE, Pouget A. Neural correlations, population coding and computation. Nat Rev Neurosci. 2006;7:358–366. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
