

Hyperglycemia activates caspase-1 and TXNIP-mediated IL-1beta transcription in human adipose tissue
- PMID: 21270263
- PMCID: PMC3028351
- DOI: 10.2337/db10-0266
Hyperglycemia activates caspase-1 and TXNIP-mediated IL-1beta transcription in human adipose tissue
Abstract
Objective: Obesity is characterized by elevated levels of proinflammatory cytokines, including interleukin (IL)-1β, that contribute to the development of insulin resistance. In this study, we set out to investigate whether hyperglycemia drives IL-1β production and caspase-1 activation in murine and human adipose tissue, thus inducing insulin resistance.
Research design and methods: ob/ob animals were used as a model to study obesity and hyperglycemia. Human adipose tissue fragments or adipocytes were cultured in medium containing normal or high glucose levels. Additionally, the role of thioredoxin interacting protein (TXNIP) in glucose-induced IL-1β production was assessed.
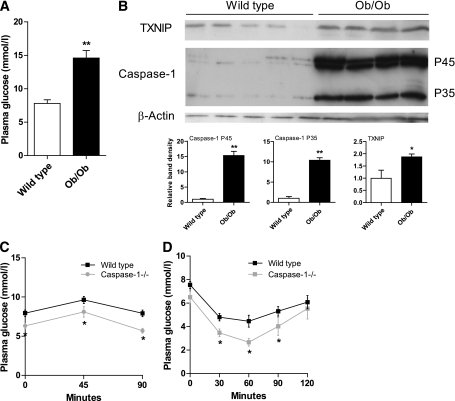
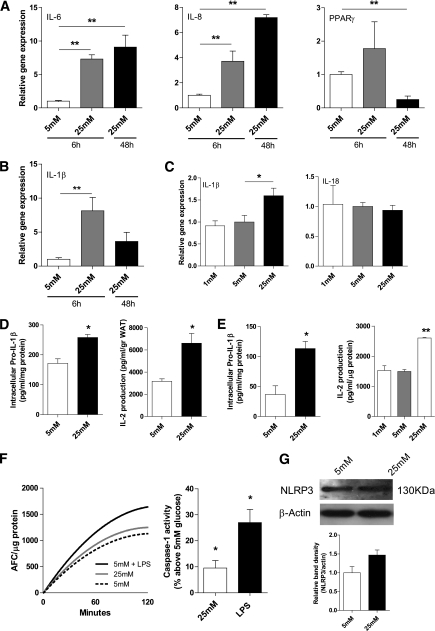
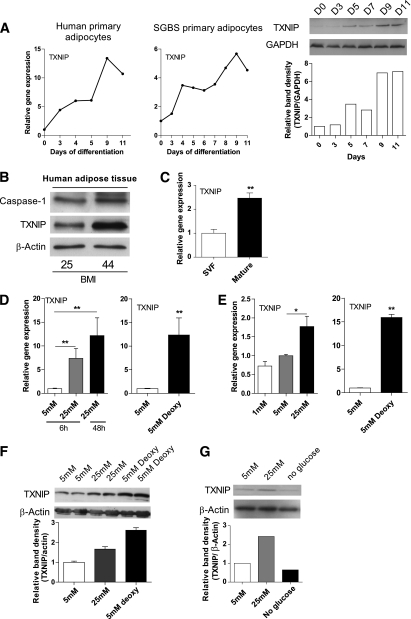
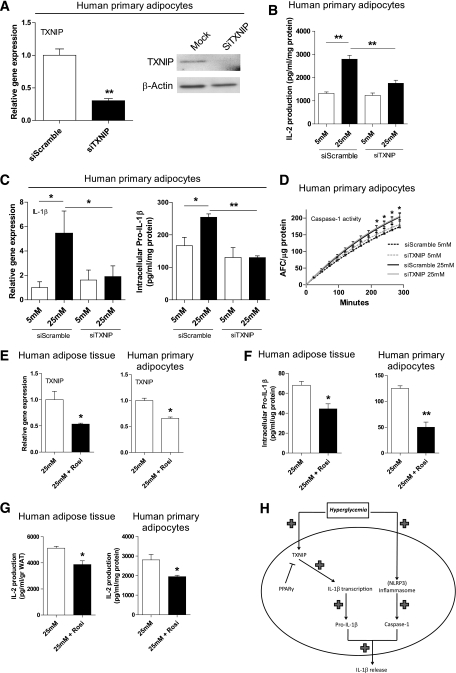
Results: TXNIP and caspase-1 protein levels were more abundantly expressed in adipose tissue of hyperglycemic ob/ob animals as compared with wild-type mice. In human adipose tissue, high glucose resulted in a 10-fold upregulation of TXNIP gene expression levels (P < 0.01) and a 10% elevation of caspase-1 activity (P < 0.05), together with induction of IL-1β transcription (twofold, P < 0.01) and a significant increase in IL-1β secretion. TXNIP suppression in human adipocytes, either by a small interfering RNA approach or a peroxisome proliferator-activated receptor-γ agonist, counteracted the effects of high glucose on bioactive IL-1 production (P < 0.01) mainly through a decrease in transcription levels paralleled by reduced intracellular pro-IL-1β levels.
Conclusions: High glucose activates caspase-1 in human and murine adipose tissue. Glucose-induced activation of TXNIP mediates IL-1β mRNA expression levels and intracellular pro-IL-1β accumulation in adipose tissue. The concerted actions lead to enhanced secretion of IL-1β in adipose tissue that may contribute to the development of insulin resistance.
Figures
References
-
- Alexandraki K, Piperi C, Kalofoutis C, Singh J, Alaveras A, Kalofoutis A. Inflammatory process in type 2 diabetes: the role of cytokines. Ann N Y Acad Sci 2006;1084:89–117 - PubMed
-
- Katsuki A, Sumida Y, Murashima S, et al. Serum levels of tumor necrosis factor-alpha are increased in obese patients with noninsulin-dependent diabetes mellitus. J Clin Endocrinol Metab 1998;83:859–862 - PubMed
-
- Shoelson SE, Herrero L, Naaz A. Obesity, inflammation, and insulin resistance. Gastroenterology 2007;132:2169–2180 - PubMed
-
- Lin Y, Rajala MW, Berger JP, Moller DE, Barzilai N, Scherer PE. Hyperglycemia-induced production of acute phase reactants in adipose tissue. J Biol Chem 2001;276:42077–42083 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
