

Transient Hoogsteen base pairs in canonical duplex DNA
- PMID: 21270796
- PMCID: PMC3074620
- DOI: 10.1038/nature09775
Transient Hoogsteen base pairs in canonical duplex DNA
Abstract
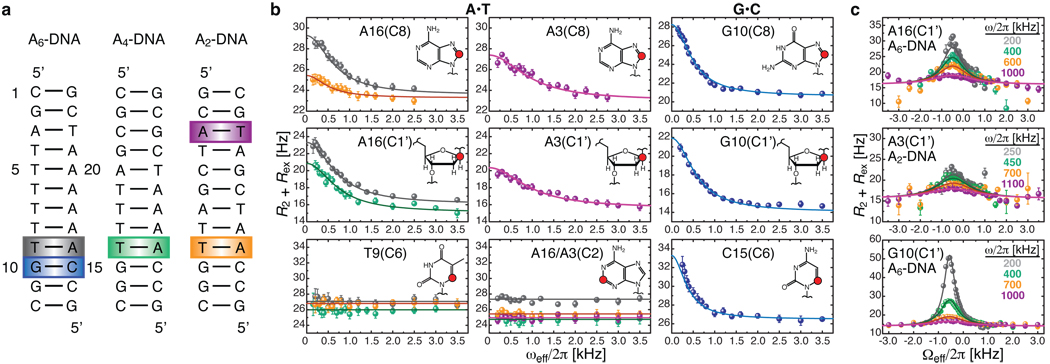
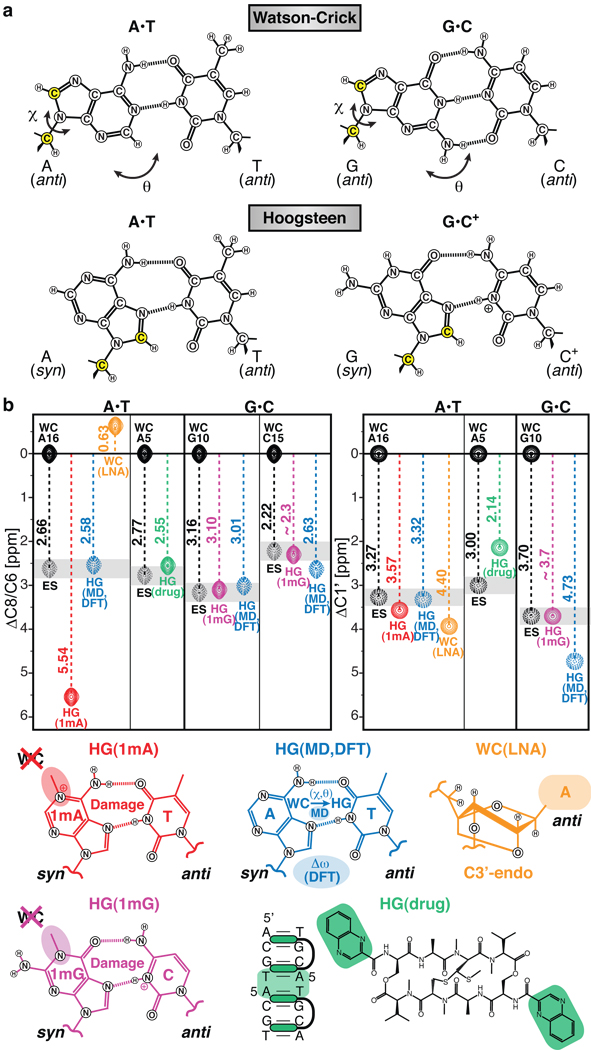
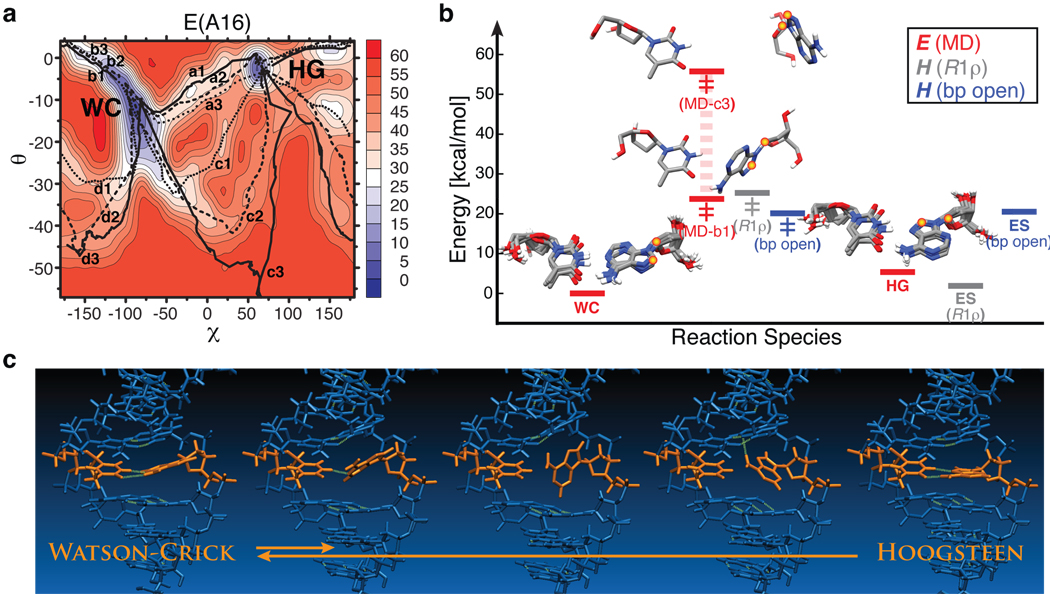
Sequence-directed variations in the canonical DNA double helix structure that retain Watson-Crick base-pairing have important roles in DNA recognition, topology and nucleosome positioning. By using nuclear magnetic resonance relaxation dispersion spectroscopy in concert with steered molecular dynamics simulations, we have observed transient sequence-specific excursions away from Watson-Crick base-pairing at CA and TA steps inside canonical duplex DNA towards low-populated and short-lived A•T and G•C Hoogsteen base pairs. The observation of Hoogsteen base pairs in DNA duplexes specifically bound to transcription factors and in damaged DNA sites implies that the DNA double helix intrinsically codes for excited state Hoogsteen base pairs as a means of expanding its structural complexity beyond that which can be achieved based on Watson-Crick base-pairing. The methods presented here provide a new route for characterizing transient low-populated nucleic acid structures, which we predict will be abundant in the genome and constitute a second transient layer of the genetic code.
Figures

Comment in
-
Biophysics: Flipping Watson and Crick.Nature. 2011 Feb 24;470(7335):472-3. doi: 10.1038/470472a. Nature. 2011. PMID: 21350476 No abstract available.
References
-
- Watson JD, Crick FH. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid. Nature. 1953;171:737–738. - PubMed
-
- Record MT, Jr, et al. Double helical DNA: conformations, physical properties, and interactions with ligands. Annu Rev Biochem. 1981;50:997–1024. - PubMed
-
- Koudelka GB, Mauro SA, Ciubotaru M. Indirect readout of DNA sequence by proteins: the roles of DNA sequence-dependent intrinsic and extrinsic forces. Prog Nucleic Acid Res Mol Biol. 2006;81:143–177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
