

Review
doi: 10.1152/jn.00960.2010.
Epub 2011 Jan 27.
Nonlinear effects of noradrenergic modulation of olfactory bulb function in adult rodents
Affiliations
- PMID: 21273323
- PMCID: PMC3075300
- DOI: 10.1152/jn.00960.2010
Item in Clipboard
Review
Nonlinear effects of noradrenergic modulation of olfactory bulb function in adult rodents
J Neurophysiol.
2011 Apr.
Abstract
The mammalian main olfactory bulb receives a significant noradrenergic input from the locus coeruleus. Norepinephrine (NE) is involved in acquisition of conditioned odor preferences in neonatal animals, in some species-specific odor-dependent behaviors, and in adult odor perception. We provide a detailed review of the functional role of NE in adult rodent main olfactory bulb function. We include cellular, synaptic, network, and behavioral data and use computational simulations to tie these different types of data together.
Figures
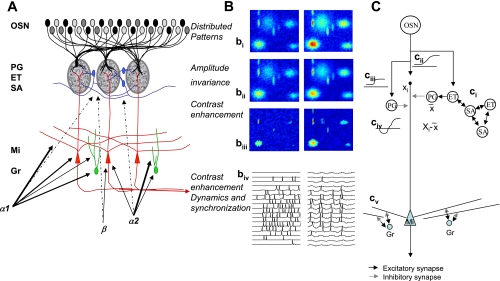
Olfactory bulb processing. A: schematic of olfactory bulb organization. Olfactory sensory neurons (OSN) expressing a common receptor, and therefore exhibiting similar odorant receptive fields, project to 2 common glomeruli in the main olfactory bulb (MOB). Within this glomerulus, OSNs make excitatory synapses onto mitral and tufted cells and the primary output neurons of the MOB, as well as glomerular layer interneurons comprising periglomerular (PG) and external tufted (ET) cells. Most PG cells (∼70%) and a third type of interneuron, short axon cells (SA), are not directly activated by OSNs. PG, ET, and SA cells form intricate networks within the glomerular layer that have been proposed to perform operations such as normalization, contrast enhancement, and synchronization. In deeper processing layers, mitral cells (Mi) interact with at least 1 other class of interneurons, granule cells (Gr). These provide extensive feedback and lateral interactions between Mi cells by interacting with their elongated secondary dendrites. This layer of processing is thought to be involved in creating olfactory bulb gamma rhythms and generating synchronized spike patterns. Noradrenergic inputs from the locus coeruleus activate 3 classes of noradrenergic receptors distributed across the MOB. α1-Receptors are thought to be predominantly located on Mi and Gr cell bodies, as well as secondary dendrites, with a sparser distribution in the glomerular layers; α2-receptors are mainly located on granule cells with a sparse distribution on Mi cell bodies and the glomerular layer; and β-receptors have been reported on Mi cell bodies and in the glomerular layer. B: schematic of glomerular input layer and Mi cell activity patterns in response to odor stimulation. bi-–biii show simulated distributed odor responses at 2 concentrations, with the lower concentration at left and high concentration at right. The 2-dimensional simulations are color coded, with red indicating high, and dark blue low, levels of activity. bii shows the same patterns after amplitude invariance processing has been performed by the network of local interneurons, and biii shows the same pattern after contrast enhancement. The patterns in biii represent the end result of the glomerular computations transmitted to deeper layers by Mi cells. The details of these computations are given by Cleland et al. (2007) and Cleland and Sethupathy (2006). biv shows how spikes generated in Mi cells in response to activation patterns (left) are transformed into sparser, oscillatory and highly synchronized spike patterns (right) by the interactions with deeper interneuron networks (Escanilla et al. 2010; Mandairon et al. 2006b). C: neural networks that could perform bulbar computations. ci indicates amplitude invariance computation as proposed by Cleland et al. (2007). ET cells sample OSN activity across the glomerular layer. A localized network of SA and ET cells, densely interconnected, calculates the sum or average activity amplitude (X̄) across the glomerular layer and projects this average activity onto inhibitory PG cells. PG cells inhibit Mi cells, hence Mi cells, which sample a single data point Xi from OSN, inputs process Xi − X̄, corresponding to a z-score normalization. cii–civ: local contrast enhancement as proposed by Cleland and Sethupathy (2006). OSN odor responses (cii), schematically drawn as response amplitude as a function of receptor-odorant affinity, activate Mi cells and PG cells. Because of the higher input resistance of PG cells, the corresponding response curves are steeper and more easily saturated (ciii). Because PG cells locally inhibit Mi cells, the resulting net effect on Mi cells is a Mexican hat function (civ). Contrast enhancement and amplitude invariance calculations are performed simultaneously and partially by the same neurons; the result is an activity pattern with higher contrast and relative amplitude invariance, as depicted in bi–biii. This activity is transformed again in deeper layers by dynamic interactions with Gr cells. Current thinking assumes that in this layer, activity patterns conveyed as Poisson-distributed spike trains (biv, left) are modulated and a fast oscillation in the gamma range is superposed on these spike trains, modulating them in time and creating periodically synchronized patterns as depicted schematically in biv (right). cv: interactions between Mi cell secondary dendrites and Gr cells provide feedback (within the same column) and lateral (between columns) inhibitory interactions. These have been proposed to create oscillatory dynamics, synchronization, and/or contrast enhancement.
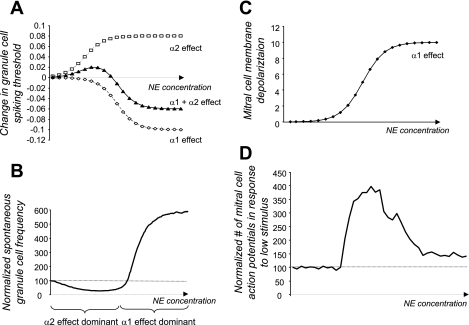
Effects of norepinephrine (NE) modulation on cellular parameters. A: NE modulates Gr cell spontaneous firing (Nai et al. 2010). The measured effects of NE activation of α1- and α2-receptors on Gr cell-mediated spontaneous inhibitory postsynaptic currents (IPSCs) was simulated by changes in Gr cell firing thresholds [in abstract parameters; equations described in detail by Escanilla et al. (2010)]. α1- and α2-receptor activation modulate spontaneous IPSCs in opposite directions, each with their own concentration dependency. The summated effect is a nonmonotonic modulation of firing threshold. B: effect of threshold modulation on Gr cell spontaneous firing. The size of the effect was adjusted to data reported by Ennis and colleagues (Nai et al. 2009, 2010). C: modulation of membrane potential by NE in model Mi cells. The size of the effect was modeled after data reported by Hayar et al. (2001). In the model, spontaneous activity was not effected by this effect, but responses to low-threshold stimuli were modulated. D: modulation of response probability to low-threshold stimuli in Mi cells. Both modulation of inhibition by α2 and α2 effects on Gr cell firing and direct modulation of Mi cell membrane potential were taken into account. Mi cell responses to low-amplitude stimuli were modulated in a nonmonotonic manner by simulated NE due to the additive effects of NE receptor activation.
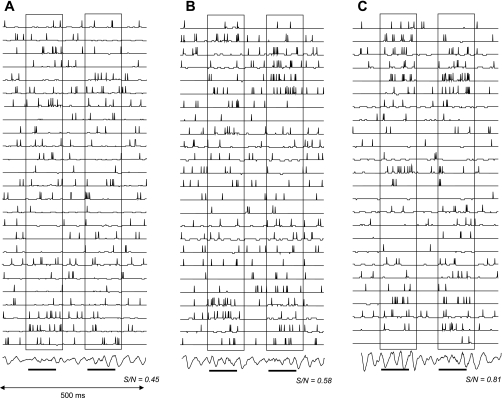
Modulation of model odor responses. Membrane potential and action potentials in a subset of olfactory bulb model Mi cells are shown in response to very low concentration odor stimuli; odor presentation is indicated by lower horizontal bars and is highlighted by outlined boxes. Odor stimulation activated OSNs to 10% of their maximal activation level. The model is modulated with no (A), low (B; mainly α2 effects), and high levels of NE (C; mainly α1 effects). The NE modulation corresponds to the negative and positive peaks of the modulation function in Fig. 2A. Signal-to-noise ratio (S/N) for each simulation is indicated below the traces and is calculated as the number of spikes evoked during odor stimulation divided by the total number of spikes evoked during the 500-ms simulation. Note that Mi cell odor responses are enhanced with respect to background activity when low NE levels are simulated and S/N is increased. At high NE levels, Mi cell responses to odorants are further increased on a background of lower spontaneous activity, and S/N further increases. Note that although the total number of spikes over the course of a 500-ms simulation does not substantially change, the distribution of spikes during spontaneous and odor activity does change. The traces at bottom represent model field potentials recorded in the Gr cell layer.

Detection and discrimination of very low concentration stimuli in model. A–C: NE modulation was simulated by varying the spiking threshold of Gr cells and Mi cell depolarization according to the sigmoid functions shown in Fig. 2. Odors were modeled as very low concentration stimuli that activated ∼30% of the available OSN population to varying degrees but to maximally 10% of their possible response magnitude. Details of OSN and odor simulations can be found in Escanilla et al. (2010) and Mandairon et al. (2006b). A: magnitude of odor detection in model neurons as a function of NE modulation. Population responses of model neurons to a simulated complex odor were compared with spontaneous activity by calculating the normalized Euclidean distance between the firing rate activation patterns [see for modeling details Escanilla et al. (2010) and Mandairon et al. (2006b)]. Odor detection in the model varied in a nonmonotonic fashion, roughly replicating that of Mi cell responsiveness depicted in Fig. 2C. B: magnitude of odor discrimination in model neurons as a function of NE modulation in the model. The magnitude of discrimination was computed as the normalized Euclidean distance between the population responses to 2 highly similar odorants. Odor discrimination increased as a function of NE modulation at medium to high NE concentrations. C: effect of NE modulation on only Mi cells in the model. If only Mi cells are affected by NE in the model, odor detection increases at the expense of discrimination. D: synchronization index (number of synchronous spikes/total number of spikes during odor response) as a function of NE modulation.

Summary of behavioral results. Behavioral results are reanalyzed from data previously published by Escanilla et al. (2010) and Mandairon et al. (2006b). A: rats with cannulas for drug infusions in both MOBs were tested on odor detection and discrimination using a habituation ask. Odors were presented in the home cage using a weighing dish with odor impregnated filter paper. B: rats were first habituated to the behavioral context by 3 successive presentations of mineral only (MO). Odor detection was then tested by presenting a novel odorant (O1; detection). After 3 habituation trials to this odorant, discrimination was tested by a single presentation of a second, chemically and perceptually similar odorant (O2). C and D: behavioral results. Rats were originally tested at a range of odor concentrations with vapor partial pressures varying from 10−2 to 10−6 Pa. Data presented pertain to the lowest concentration used only. Rats were infused with a range of NE concentrations as shown. C: odor detection magnitude at the lowest odor concentration tested (10−6 Pa) as a function of NE concentration. Detection magnitude is calculated as (O1 − H4)/(O1 + H4), where O1 is the first response to the novel odor and H4 is the response during the last habituation trial to mineral oil. This index varies between 0 (O1 = H4, no detection) and 1 (O1 >> H4, high detection) and is normalized with respect to individual differences in sniff times. Note that detection magnitude follows a nonmonotonic function of NE concentration, as would be predicted by the nonmonotonic dependency of cellular modulation by NE. D: odor discrimination magnitude at the lowest odor concentration tested (10−6 Pa) as a function of NE concentration. Discrimination magnitude is calculated as (O2 − O1)/(O2 + O1), where O2 is the response to the second odor and O1 is the response during the last trial with O1. The index varies between 0 (O2 = O1, no discrimination) and 1 (O2 >> O1, high discrimination) and is normalized with respect to individual sniff times.

Summary of NE effects in the OB. Activation of α1-receptors depolarizes Mi cells and increases spontaneous activity in Gr cell. The net result of these actions is an increase in excitability accompanied by an increase in oscillatory power. Behaviorally, this results in lower detection and discrimination thresholds.
References
-
- Aston-Jones G, Rajkowski J, Cohen J. Locus coeruleus and regulation of behavioral flexibility and attention. Prog Brain Res 126: 165–182, 2000 - PubMed
-
- Aungst JL, Heyward PM, Puche AC, Karnup SV, Hayar A, Szabo G, Shipley MT. Centre-surround inhibition among olfactory bulb glomeruli. Nature 426: 623–629, 2003 - PubMed
-
- Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev 42: 33–84, 2003 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous