

Claudins: control of barrier function and regulation in response to oxidant stress
- PMID: 21275791
- PMCID: PMC3144428
- DOI: 10.1089/ars.2011.3893
Claudins: control of barrier function and regulation in response to oxidant stress
Abstract
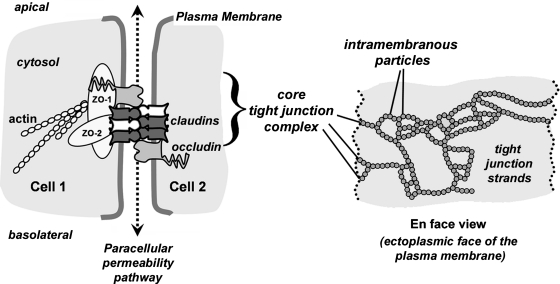
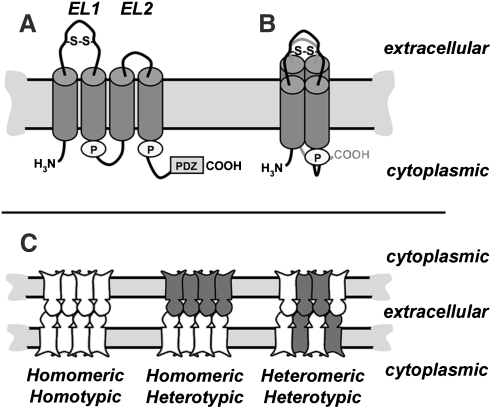
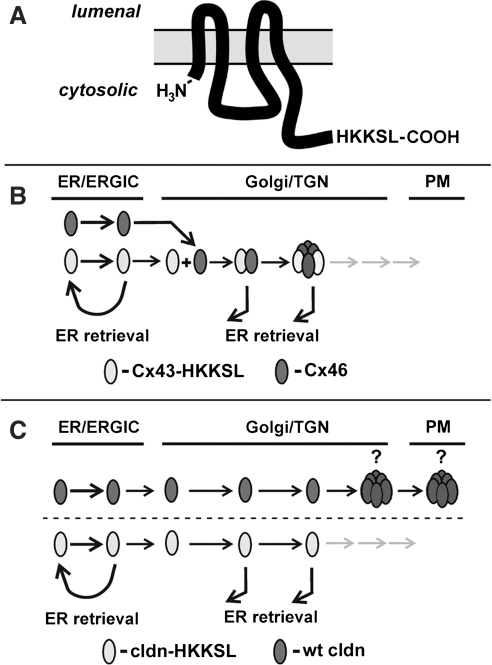
Claudins are a family of nearly two dozen transmembrane proteins that are a key part of the tight junction barrier that regulates solute movement across polarized epithelia. Claudin family members interact with each other, as well as with other transmembrane tight junction proteins (such as occludin) and cytosolic scaffolding proteins (such as zonula occludens-1 (ZO-1)). Although the interplay between all of these different classes of proteins is critical for tight junction formation and function, claudin family proteins are directly responsible for forming the equivalent of paracellular ion selective channels (or pores) with specific permeability and thus are essential for barrier function. In this review, we summarize current progress in identifying structural elements of claudins that regulate their transport, assembly, and function. The effects of oxidant stress on claudins are also examined, with particular emphasis on lung epithelial barrier function and oxidant stress induced by chronic alcohol abuse.
Figures



References
-
- Amasheh S. Meiri N. Gitter AH. Schoneberg T. Mankertz J. Schulzke JD. Fromm M. Claudin-2 expression induces cation-selective channels in tight junctions of epithelial cells. J Cell Sci. 2002;115:4969–4976. - PubMed
-
- Amasheh S. Schmidt T. Mahn M. Florian P. Mankertz J. Tavalali S. Gitter AH. Schulzke JD. Fromm M. Contribution of claudin-5 to barrier properties in tight junctions of epithelial cells. Cell Tissue Res. 2005;321:89–96. - PubMed
-
- Andras IE. Pu H. Deli MA. Nath A. Hennig B. Toborek M. HIV-1 Tat protein alters tight junction protein expression and distribution in cultured brain endothelial cells. J Neurosci Res. 2003;74:255–265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
