

Glutamate pharmacology and metabolism in peripheral primary afferents: physiological and pathophysiological mechanisms
- PMID: 21276816
- PMCID: PMC5937940
- DOI: 10.1016/j.pharmthera.2011.01.005
Glutamate pharmacology and metabolism in peripheral primary afferents: physiological and pathophysiological mechanisms
Abstract
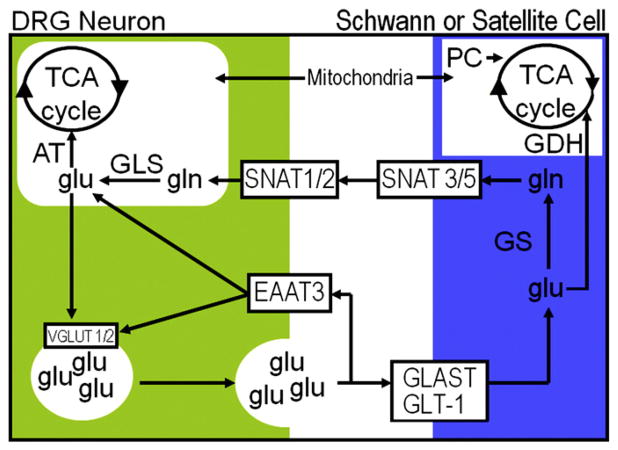
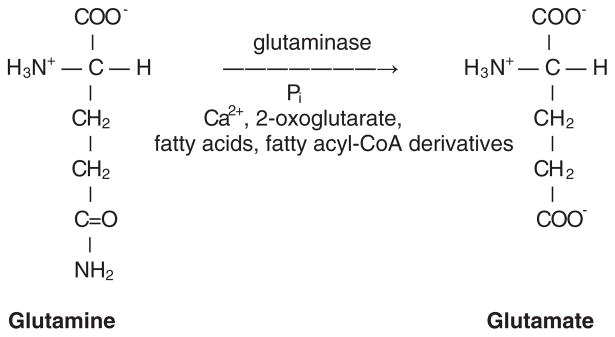
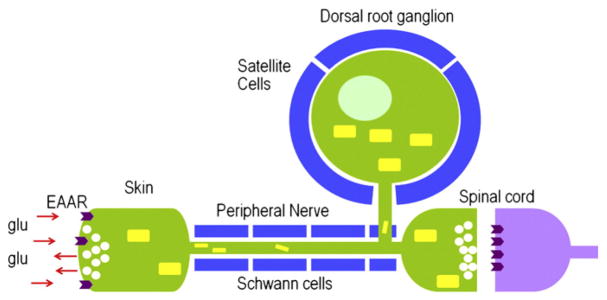
In addition to using glutamate as a neurotransmitter at central synapses, many primary sensory neurons release glutamate from peripheral terminals. Primary sensory neurons with cell bodies in dorsal root or trigeminal ganglia produce glutaminase, the synthetic enzyme for glutamate, and transport the enzyme in mitochondria to peripheral terminals. Vesicular glutamate transporters fill neurotransmitter vesicles with glutamate and they are shipped to peripheral terminals. Intense noxious stimuli or tissue damage causes glutamate to be released from peripheral afferent nerve terminals and augmented release occurs during acute and chronic inflammation. The site of action for glutamate can be at the autologous or nearby nerve terminals. Peripheral nerve terminals contain both ionotropic and metabotropic excitatory amino acid receptors (EAARs) and activation of these receptors can lower the activation threshold and increase the excitability of primary afferents. Antagonism of EAARs can reduce excitability of activated afferents and produce antinociception in many animal models of acute and chronic pain. Glutamate injected into human skin and muscle causes acute pain. Trauma in humans, such as arthritis, myalgia, and tendonitis, elevates glutamate levels in affected tissues. There is evidence that EAAR antagonism at peripheral sites can provide relief in some chronic pain sufferers.
Copyright © 2011. Published by Elsevier Inc.
Figures


Similar articles
-
Modulation of sensory input to the spinal cord by presynaptic ionotropic glutamate receptors.Arch Ital Biol. 2005 May;143(2):103-12. Arch Ital Biol. 2005. PMID: 16106991 Review.
-
Vanilloid receptor VR1-positive primary afferents are glutamatergic and contact spinal neurons that co-express neurokinin receptor NK1 and glutamate receptors.J Neurocytol. 2004 May;33(3):321-9. doi: 10.1023/B:NEUR.0000044193.31523.a1. J Neurocytol. 2004. PMID: 15475687
-
Inhibition of glutamate uptake in the spinal cord induces hyperalgesia and increased responses of spinal dorsal horn neurons to peripheral afferent stimulation.Neuroscience. 2006;138(4):1351-60. doi: 10.1016/j.neuroscience.2005.11.061. Epub 2006 Jan 19. Neuroscience. 2006. PMID: 16426766
-
Glutamate-induced sensitization of rat masseter muscle fibers.Neuroscience. 2002;109(2):389-99. doi: 10.1016/s0306-4522(01)00489-4. Neuroscience. 2002. PMID: 11801373
-
Glutamatergic functions of primary afferent neurons with special emphasis on vagal afferents.Int Rev Cytol. 2007;256:223-75. doi: 10.1016/S0074-7696(07)56007-9. Int Rev Cytol. 2007. PMID: 17241909 Review.
Cited by
-
Topical versus caudal ketamine/bupivacaine combination for postoperative analgesia in children undergoing inguinal herniotomy.Saudi J Anaesth. 2017 Jan-Mar;11(1):41-48. doi: 10.4103/1658-354X.197338. Saudi J Anaesth. 2017. PMID: 28217052 Free PMC article.
-
Tumour-Derived Glutamate: Linking Aberrant Cancer Cell Metabolism to Peripheral Sensory Pain Pathways.Curr Neuropharmacol. 2017;15(4):620-636. doi: 10.2174/1570159X14666160509123042. Curr Neuropharmacol. 2017. PMID: 27157265 Free PMC article.
-
Sonic Hedgehog Signaling Pathway: A Role in Pain Processing.Neurochem Res. 2023 Jun;48(6):1611-1630. doi: 10.1007/s11064-023-03864-5. Epub 2023 Feb 4. Neurochem Res. 2023. PMID: 36738366 Review.
-
Large conductance calcium-activated potassium channels: their expression and modulation of glutamate release from nerve terminals isolated from rat trigeminal caudal nucleus and cerebral cortex.Neurochem Res. 2014 May;39(5):901-10. doi: 10.1007/s11064-014-1287-1. Epub 2014 Mar 26. Neurochem Res. 2014. PMID: 24667981
-
Chronic musculoskeletal pain: review of mechanisms and biochemical biomarkers as assessed by the microdialysis technique.J Pain Res. 2014 Jun 12;7:313-26. doi: 10.2147/JPR.S59144. eCollection 2014. J Pain Res. 2014. PMID: 24966693 Free PMC article. Review.
References
-
- Ahn DK, Jung CY, Lee HJ, Choi HS, Ju JS, Bae YC. Peripheral glutamate receptors participate in interleukin-1beta-induced mechanical allodynia in the orofacial area of rats. Neurosci Lett. 2004;357(3):203–206. - PubMed
-
- Ahn DK, Kim KH, Jung CY, Choi HS, Lim EJ, Youn DH, et al. Role of peripheral group I and II metabotropic glutamate receptors in IL-1beta-induced mechanical allodynia in the orofacial area of conscious rats. Pain. 2005;118(1–2):53–60. - PubMed
-
- Aley KO, Levine JD. Different peripheral mechanisms mediate enhanced nociception in metabolic/toxic and traumatic painful peripheral neuropathies in the rat. Neuroscience. 2002;111(2):389–397. - PubMed
-
- Alfredson H, Lorentzon R. Chronic tendon pain: no signs of chemical inflammation but high concentrations of the neurotransmitter glutamate. Implications for treatment? Curr Drug Targets. 2002;3(1):43–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
