

Complex primary afferents: What the distribution of electrophysiologically-relevant phenotypes within the spiral ganglion tells us about peripheral neural coding
- PMID: 21276843
- PMCID: PMC3223913
- DOI: 10.1016/j.heares.2011.01.014
Complex primary afferents: What the distribution of electrophysiologically-relevant phenotypes within the spiral ganglion tells us about peripheral neural coding
Abstract
Spiral ganglion neurons are the first neural element of the auditory system. They receive precise synaptic signals which represent features of sound stimuli encoded by hair cell receptors and they deliver a digital representation of this information to the central nervous system. It is well known that spiral ganglion neurons are selectively responsive to specific sound frequencies, and that numerous structural and physiological specializations in the inner ear increase the quality of this tuning, beyond what could be accomplished by the passive properties of the basilar membrane. Further, consistent with what we know about other sensory systems, it is becoming clear that the parallel divergent innervation pattern of type I spiral ganglion neurons has the potential to encode additional features of sound stimuli. To date, we understand the most about the sub-modalities of frequency and intensity coding in the peripheral auditory system. Work reviewed herein will address the issue of how intrinsic electrophysiological features of the neurons themselves have the potential to contribute to the precision of coding and transmitting information about these two parameters to higher auditory centers for further processing.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
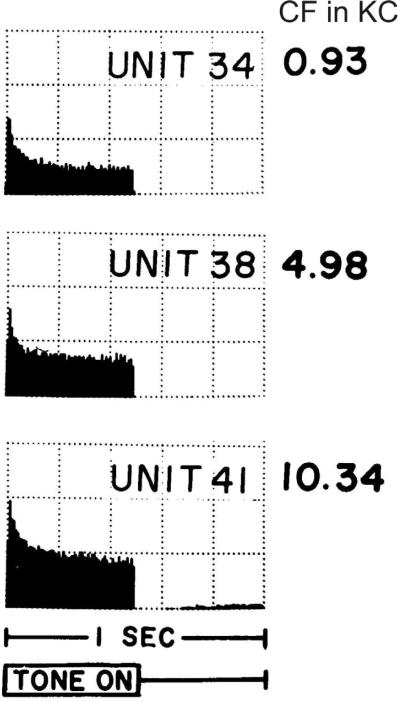
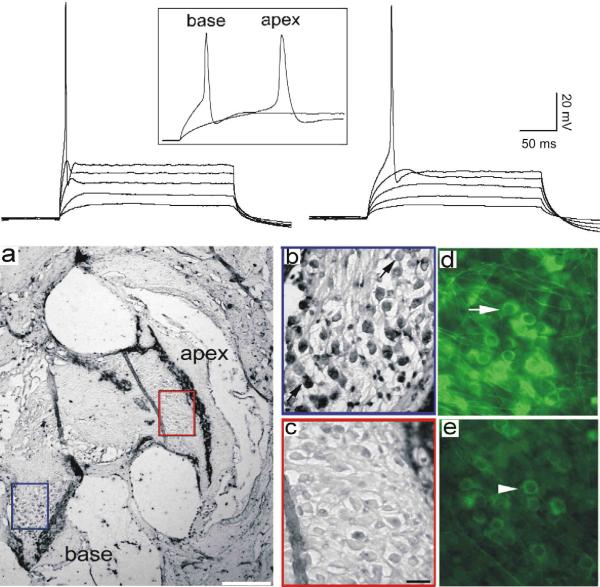
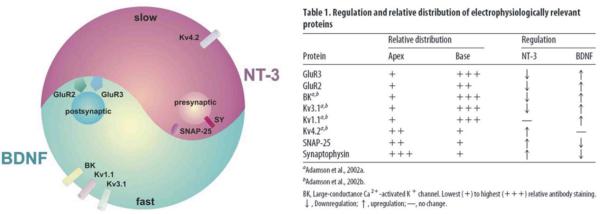
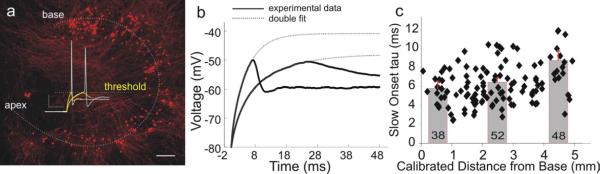
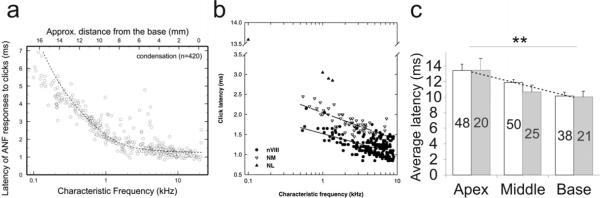
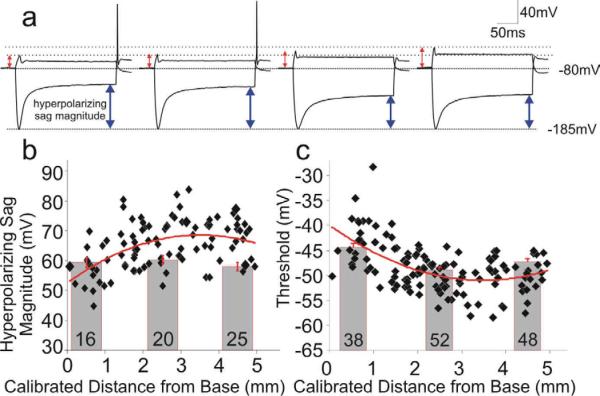
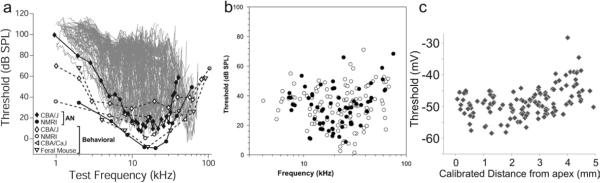
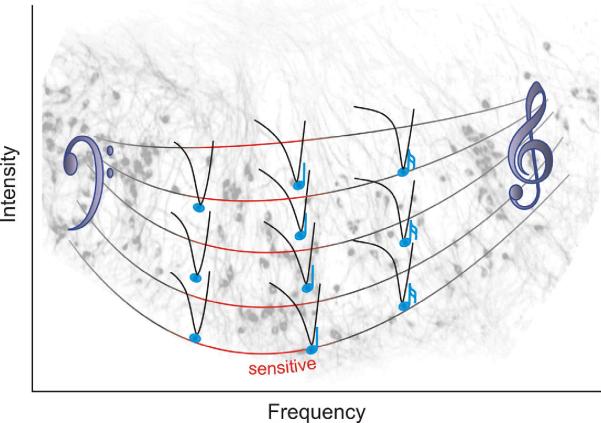
References
-
- Adamson CL, Reid MA, Mo ZL, Bowne-English J, Davis RL. Firing features and potassium channel content of murine spiral ganglion neurons vary with cochlear location. J Comp Neurol. 2002b;447:331–50. - PubMed
-
- Anniko M, Arnold W, Stigbrand T, Strom A. The human spiral ganglion. ORL J Otorhinolaryngol Relat Spec. 1995;57:68–77. - PubMed
-
- Ashmore J, Gale J. The cochlea. Curr Biol. 2000;10:R325–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
