

The Three Musketeers of Autophagy: phosphorylation, ubiquitylation and acetylation
- PMID: 21277210
- PMCID: PMC3714536
- DOI: 10.1016/j.tcb.2010.12.006
The Three Musketeers of Autophagy: phosphorylation, ubiquitylation and acetylation
Abstract
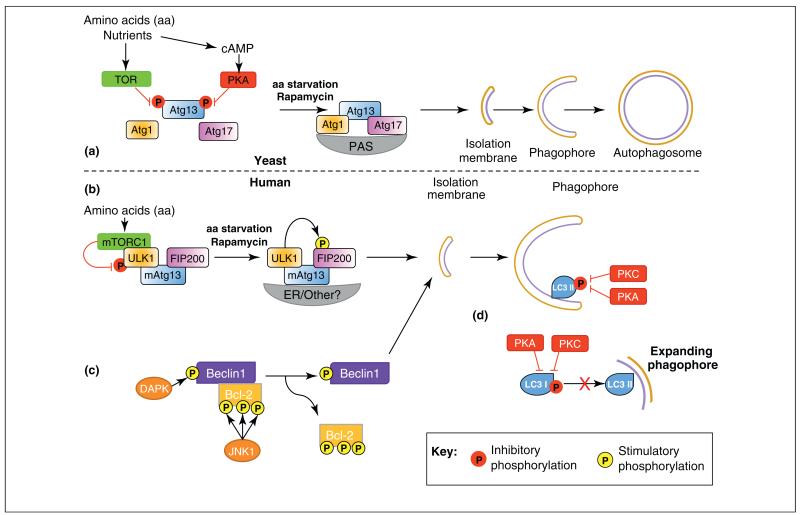
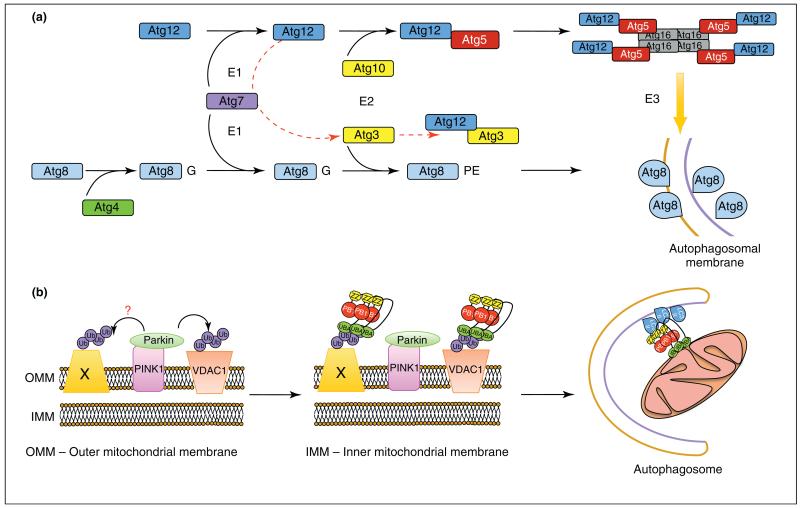
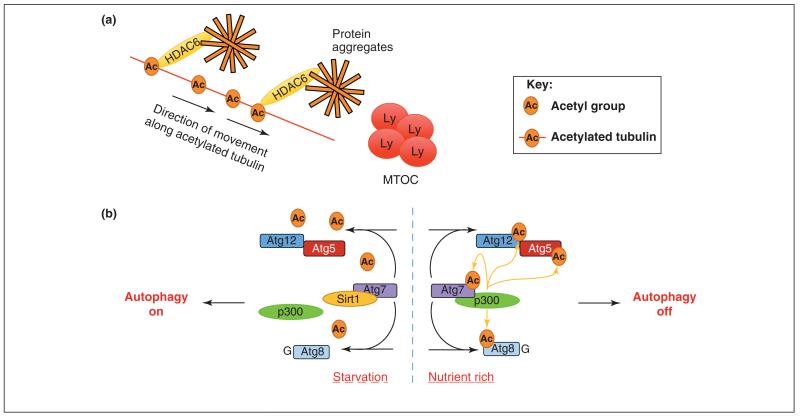
Autophagy is a highly conserved process that allows cells, tissues and organs to survive onslaughts such as nutrient deprivation, inflammation, hypoxia and other stresses. The core component proteins that regulate autophagy are well known, and the formation of a double-membrane structure that encompasses cytosolic cargo, including protein aggregates and organelles, has been intensively studied. However, less is known about the inputs that specifically alter recruitment of these components and how post-translational modifications can influence autophagy flux, or the rate at which autophagy substrates are turned over. We propose that three types of post-translational modifications - phosphorylation, ubiquitylation and acetylation - are crucial for autophagy induction, regulation and fine-tuning, and are influenced by a variety of stimuli. Understanding these novel mechanisms of autophagy regulation will give us deeper insights into this process and potentially open up therapeutic avenues.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Hayashi-Nishino M, et al. A subdomain of the endoplasmic reticulum forms a cradle for autophagosome formation. Nat. Cell Biol. 2009;11:1433–1437. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
