

The second RNA chaperone, Hfq2, is also required for survival under stress and full virulence of Burkholderia cenocepacia J2315
- PMID: 21278292
- PMCID: PMC3067662
- DOI: 10.1128/JB.01375-10
The second RNA chaperone, Hfq2, is also required for survival under stress and full virulence of Burkholderia cenocepacia J2315
Retraction in
-
Retraction for Ramos et al., The second RNA chaperone, Hfq2, is also required for survival under stress and full virulence of Burkholderia cenocepacia J2315.J Bacteriol. 2014 Nov;196(22):3980. doi: 10.1128/JB.02242-14. J Bacteriol. 2014. PMID: 25319526 Free PMC article.
Abstract
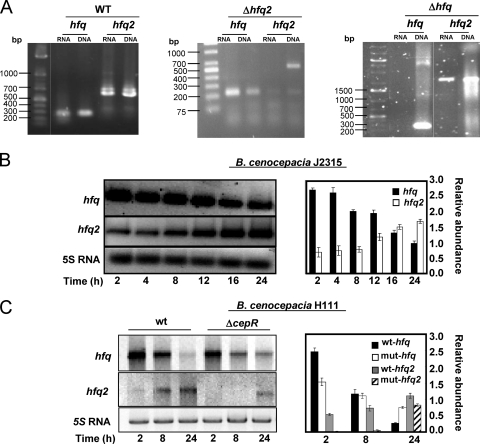
Burkholderia cenocepacia J2315 is a highly virulent and epidemic clinical isolate of the B. cepacia complex (Bcc), a group of bacteria that have emerged as important pathogens to cystic fibrosis patients. This bacterium, together with all Bcc strains and a few other prokaryotes, is unusual for encoding in its genome two distinct and functional Hfq-like proteins. In this work, we show results indicating that the 188-amino-acid Hfq2 protein is required for the full virulence and stress resistance of B. cenocepacia J2315, despite the presence on its genome of the functional 79-amino-acid Hfq protein encoded by the hfq gene. Similar to other Hfq proteins, Hfq2 is able to bind RNA. However, Hfq2 is unique in its ability to apparently form trimers in vitro. Maximal transcription of hfq was observed in B. cenocepacia J2315 cells in the early exponential phase of growth. In contrast, hfq2 transcription reached maximal levels in cells in the stationary phase, depending on the CepR quorum-sensing regulator. These results suggest that tight regulation of the expression of these two RNA chaperones is required to maximize the fitness and virulence of this bacterium. In addition, the ability of Hfq2 to bind DNA, not observed for Hfq, suggests that Hfq2 might play additional roles besides acting as an RNA chaperone.
Figures








References
-
- Andrade, M. A., P. Chacón, J. J. Merelo, and F. Morán. 1993. Evaluation of secondary structure of proteins from UV circular dichroism using an unsupervised learning neural network. Protein Eng. 6:383-390. - PubMed
-
- Brahms, S., and J. Brahms. 1980. Determination of protein secondary structure in solution by vacuum ultraviolet circular dichroism. J. Mol. Biol. 138:149-178. - PubMed
-
- Brennan, R. G., and T. M. Link. 2007. Hfq structure, function and ligand binding. Curr. Opin. Microbiol. 10:125-133. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
