

Review
doi: 10.1093/jmcb/mjq055.
BRCA1, PARP, and 53BP1: conditional synthetic lethality and synthetic viability
Affiliations
- PMID: 21278454
- PMCID: PMC3030974
- DOI: 10.1093/jmcb/mjq055
Item in Clipboard
Review
BRCA1, PARP, and 53BP1: conditional synthetic lethality and synthetic viability
J Mol Cell Biol.
2011 Feb.
Abstract
BRCA1 plays a critical role in the regulation of homologous recombination (HR)-mediated DNA double-strand break repair. BRCA1-deficient cancers have evolved to tolerate loss of BRCA1 function. This renders them vulnerable to agents, such as PARP inhibitors, that are conditionally 'synthetic lethal' with their underlying repair defect. Recent studies demonstrate that BRCA1-deficient cells may acquire resistance to these agents by partially correcting their defect in HR-mediated repair, either through reversion mutations in BRCA1 or through 'synthetic viable' loss of 53BP1. These findings and their clinical implications will be reviewed in this article.
Figures
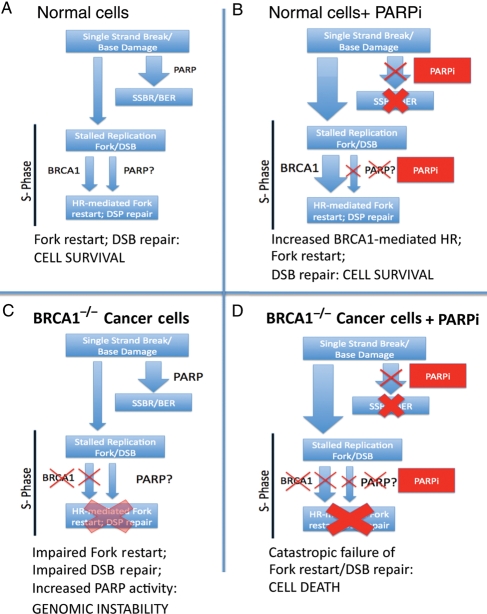
Conditional synthetic lethality of PARP inhibitors with BRCA1−/− cells. (A) In normal cells, single-strand breaks and base lesions are repaired efficiently by PARP mediated single-strand break repair (SSBR)/base excision repair (BER). The few unrepaired breaks that persist into S-phase and lead to replication fork arrest and double-strand breaks (DSBs) are efficiently handled by BRCA1-dependent HR-mediated repair and fork restart. PARP may also play a role in fork restart. (B) In the setting of PARP inhibitor (PARPi), SSBR is impaired leading to accumulation of unrepaired single-strand breaks. This results in an great increase in the amount of replication fork lesions that must be repaired by a BRCA1-dependent HR-mediated repair pathway. (C) In BRCA1−/− cells, HR mediated repair of stalled replication forks and DSBs is impaired; there may be increased reliance of PARP-dependent fork-restart mechanisms in S-phase and increased PARP activity. (D) In BRCA1−/− cells treated with PARPi, an increased burden of unrepaired single-strand breaks enter S-phase. The resulting lesions cannot be repaired, as HR-mediated repair is completely disabled, leading to catastrophic failure of replication fork restart, accumulation of unrepaired DNA DSBs and cell arrest/death.
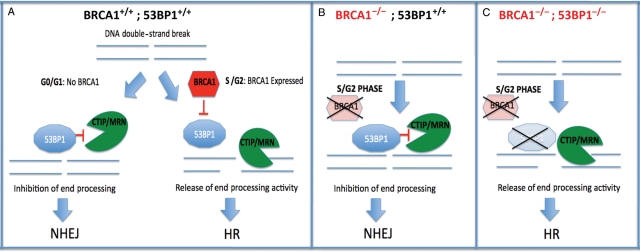
Collaboration of BRCA1 loss and 53BP1 loss results in synthetic viability and restoration of HR-mediated repair. (A) Normal cells: in G0/G1 there is no BRCA1 expression, and 53BP1 is recruited to DNA double-strand breaks (DSBs) where it restrains CtIP activity, inhibits end processing and promotes NHEJ. In S/G2 phases of the cell cycle, BRCA1 is normally expressed and recruited to sites of breaks, where it inhibits the action of 53BP1 on CtIP, allowing end processing and promoting HR mediated repair. (B) BRCA1−/− cells: BRCA1 is not present in S-phase, and 53BP1 remains free to inhibit CtIP function, leading to impaired end processing of breaks, suppression of HR and promotion of error-prone NHEJ. (C) BRCA1−/−; 53BP1−/− cells: during S-phase CtIP is recruited to sites of DNA breaks. In the absence of 53BP1, its activity is unrestrained allowing end processing and HR mediated repair.
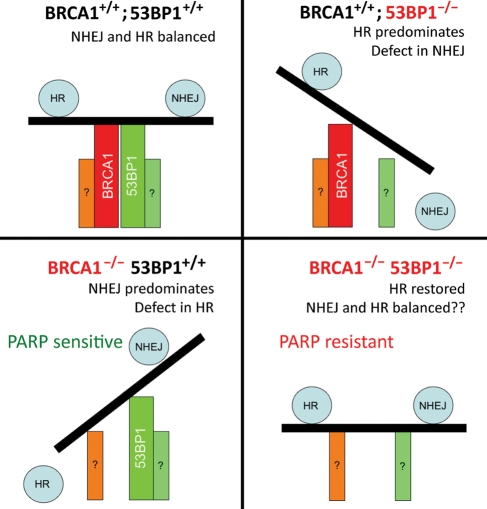
53BP1 and BRCA1 as regulators of the balance between NHEJ and HR. In normal cells (upper left), 53BP1 and BRCA1 together maintain an overall balance between NHEJ and HR. In BRCA1+/+; 53BP1−/− cells (upper right), 53BP1 loss inhibits NHEJ and promotes HR, resulting in increased frequency of HR-mediated repair. In BRCA1−/−; 53BP1+/+ cells (lower left), loss of BRCA1 results in a profound defect in HR, with 53BP1 now promoting NHEJ as the predominant repair pathway, resulting in genomic instability. In BRCA1−/−; 53BP1−/− cells (lower right), the loss of both BRCA1 and 53BP1 restores HR-capacity. The factors that now regulate choice between HR and NHEJ in this setting are unknown.
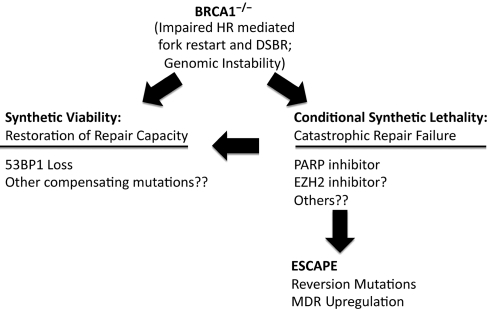
Schema showing roles of synthetic lethality, escape and synthetic viability affecting survival of BRCA1−/− cells.
References
-
- Abbott D.W., Thompson M.E., Robinson-Benion C., et al. BRCA1 expression restores radiation resistance in BRCA1-defective cancer cells through enhancement of transcription-coupled DNA repair. J. Biol. Chem. 1999;274:18808–18812. - PubMed
-
- Ahel I., Ahel D., Matsusaka T., et al. Poly(ADP-ribose)-binding zinc finger motifs in DNA repair/checkpoint proteins. Nature. 2008;451:81–85. - PubMed
-
- Allinson S.L., Dianova, Dianov G.L. Poly(ADP-ribose) polymerase in base excision repair: always engaged, but not essential for DNA damage processing. Acta Biochim. Pol. 2003;50:169–179. - PubMed
-
- Ame J.C., Rolli V., Schreiber V., et al. PARP-2, a novel mammalian DNA damage-dependent poly(ADP-ribose) polymerase. J. Biol. Chem. 1999;274:17860–17868. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
