

Fate mapping of IL-17-producing T cells in inflammatory responses
- PMID: 21278737
- PMCID: PMC3040235
- DOI: 10.1038/ni.1993
Fate mapping of IL-17-producing T cells in inflammatory responses
Abstract
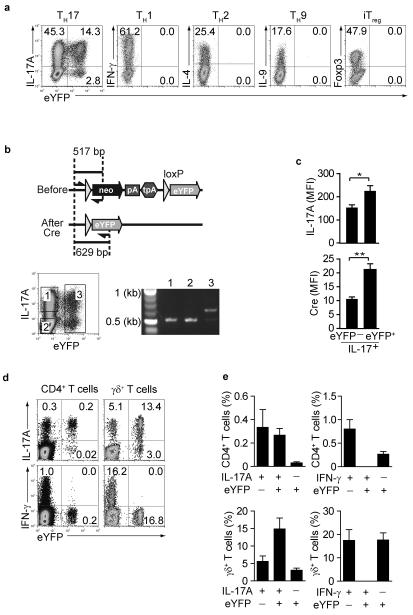
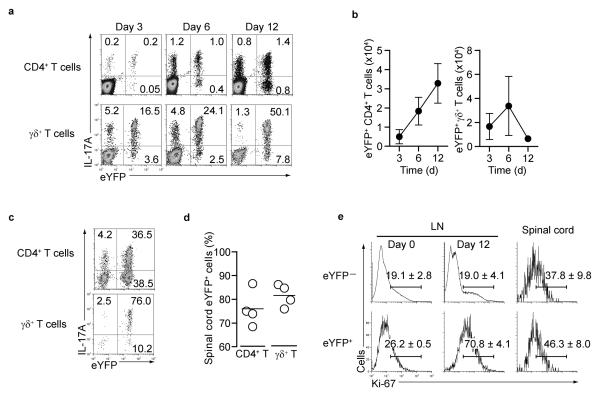
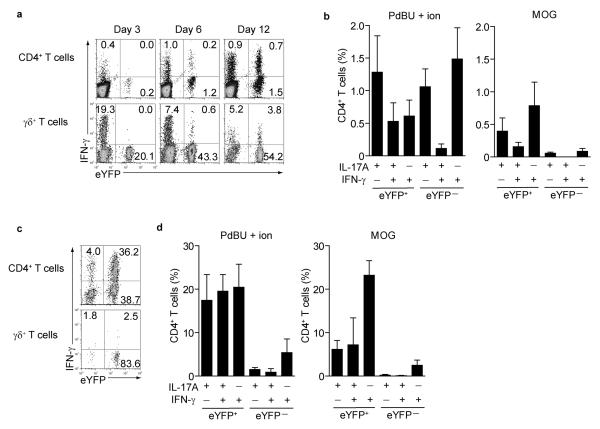
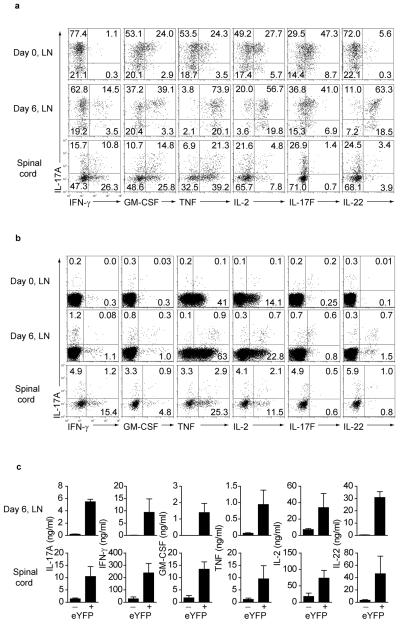
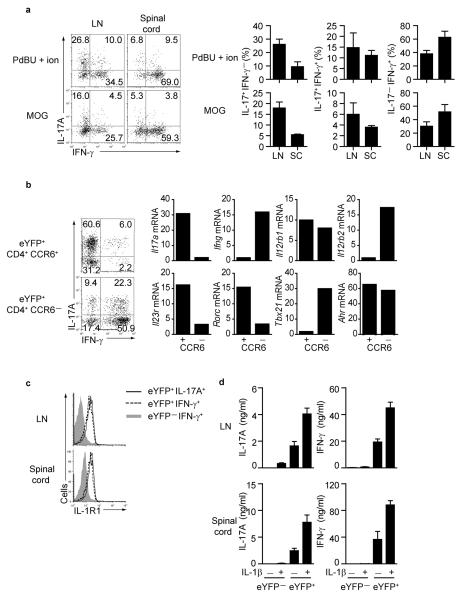
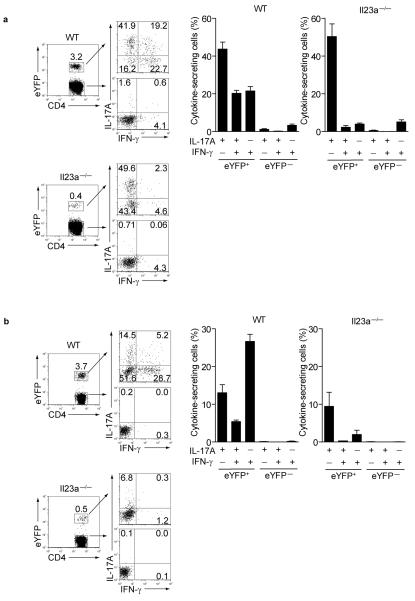
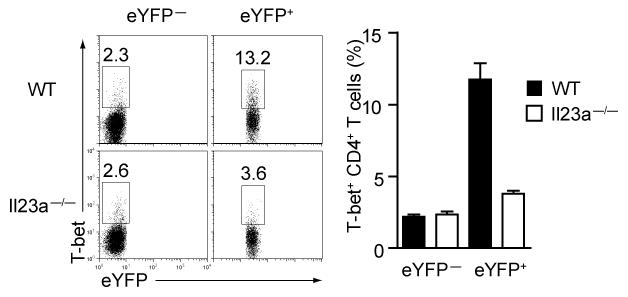
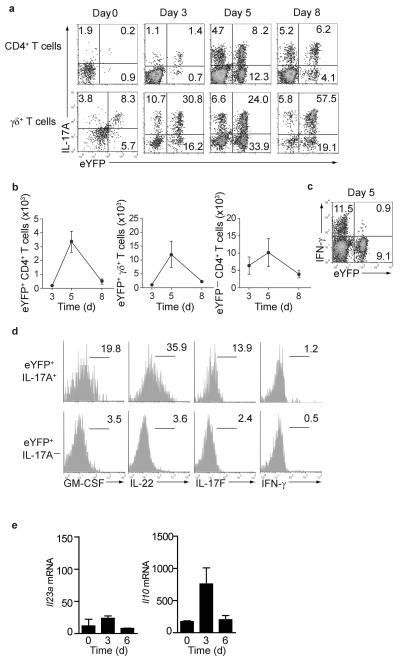
Here we describe a reporter mouse strain designed to map the fate of cells that have activated interleukin 17A (IL-17A). We found that IL-17-producing helper T cells (T(H)17 cells) had distinct plasticity in different inflammatory settings. Chronic inflammatory conditions in experimental autoimmune encephalomyelitis (EAE) caused a switch to alternative cytokines in T(H)17 cells, whereas acute cutaneous infection with Candida albicans did not result in the deviation of T(H)17 cells to the production of alternative cytokines, although IL-17A production was shut off in the course of the infection. During the development of EAE, interferon-γ (IFN-γ) and other proinflammatory cytokines in the spinal cord were produced almost exclusively by cells that had produced IL-17 before their conversion by IL-23 ('ex-T(H)17 cells'). Thus, this model allows the actual functional fate of effector T cells to be related to T(H)17 developmental origin regardless of IL-17 expression.
Figures
Comment in
-
T cells: Plastic TH17 cells.Nat Rev Immunol. 2011 Mar;11(3):160. doi: 10.1038/nri2942. Nat Rev Immunol. 2011. PMID: 21456313 No abstract available.
References
-
- Harrington LE, et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005;6(11):1123–1132. - PubMed
-
- Veldhoen M, et al. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006;24(2):179–189. - PubMed
-
- Ivanov II, et al. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell. 2006;126(6):1121–1133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
