

Competing allosteric mechanisms modulate substrate binding in a dimeric enzyme
- PMID: 21278754
- PMCID: PMC3262843
- DOI: 10.1038/nsmb.1978
Competing allosteric mechanisms modulate substrate binding in a dimeric enzyme
Abstract
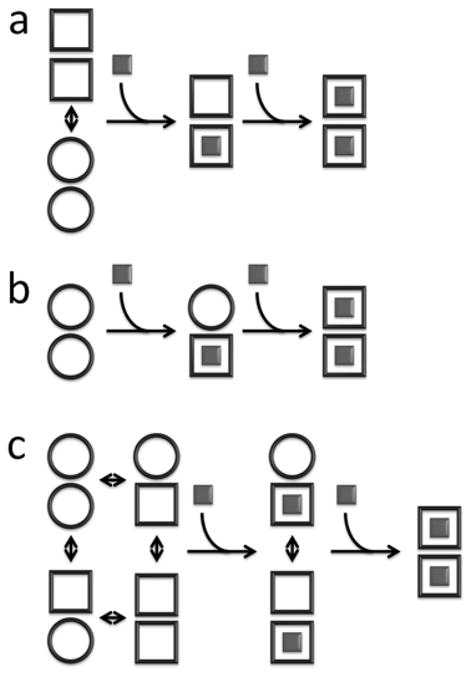
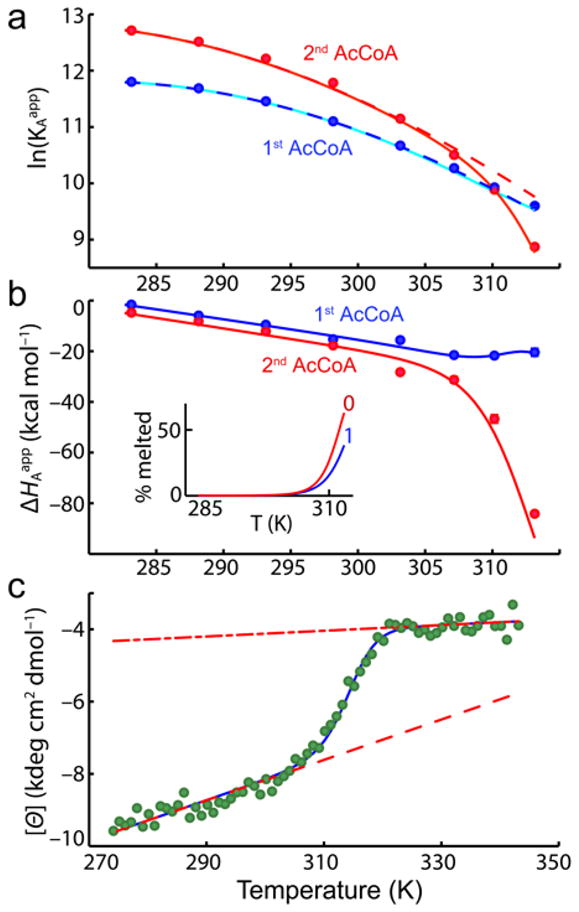
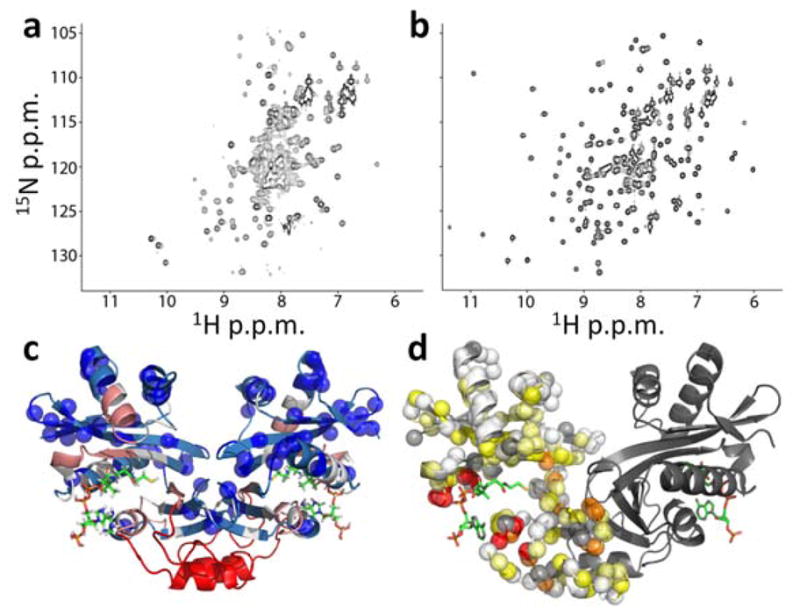
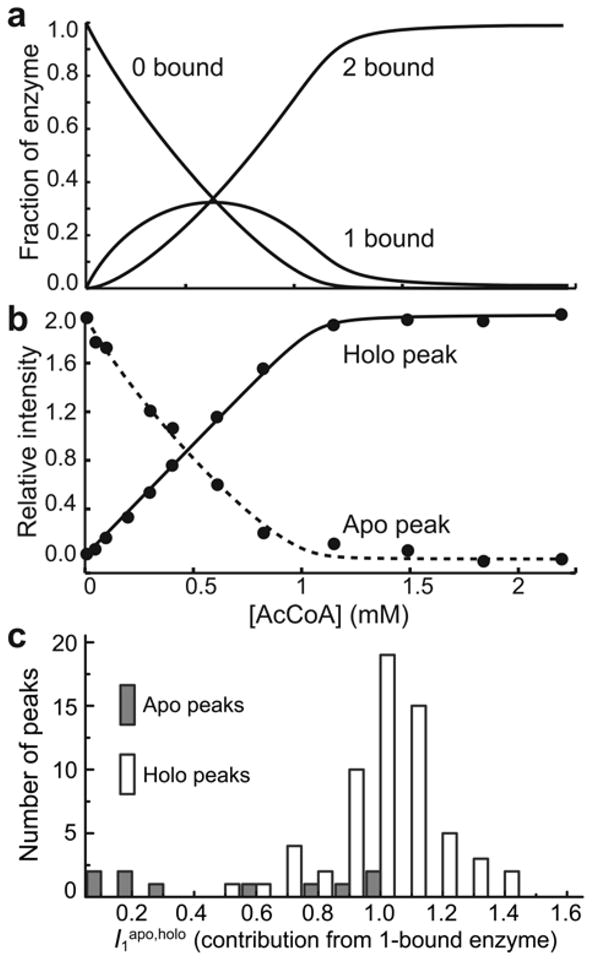
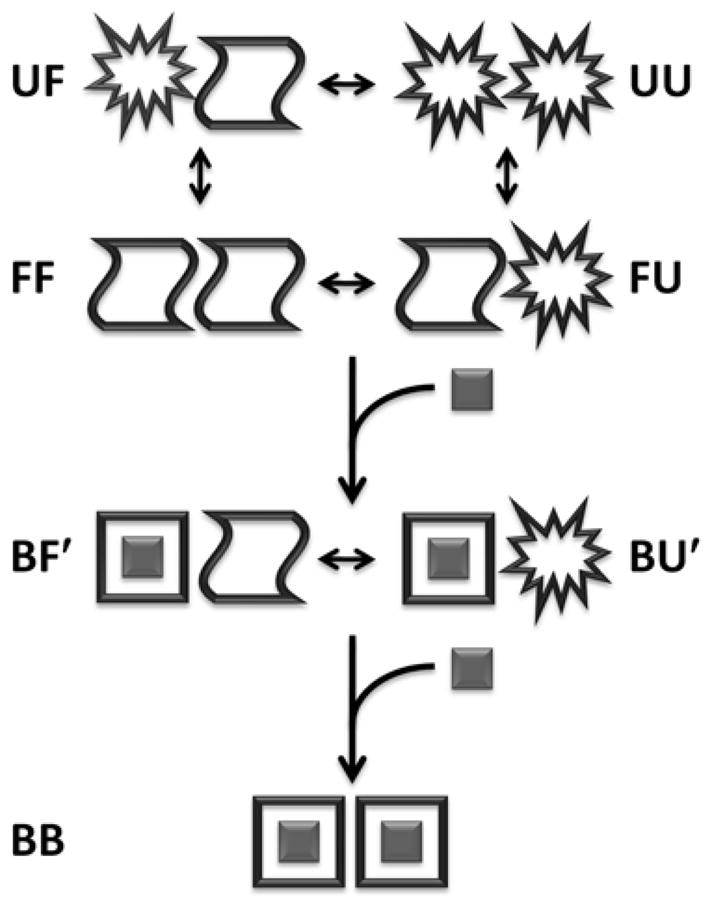
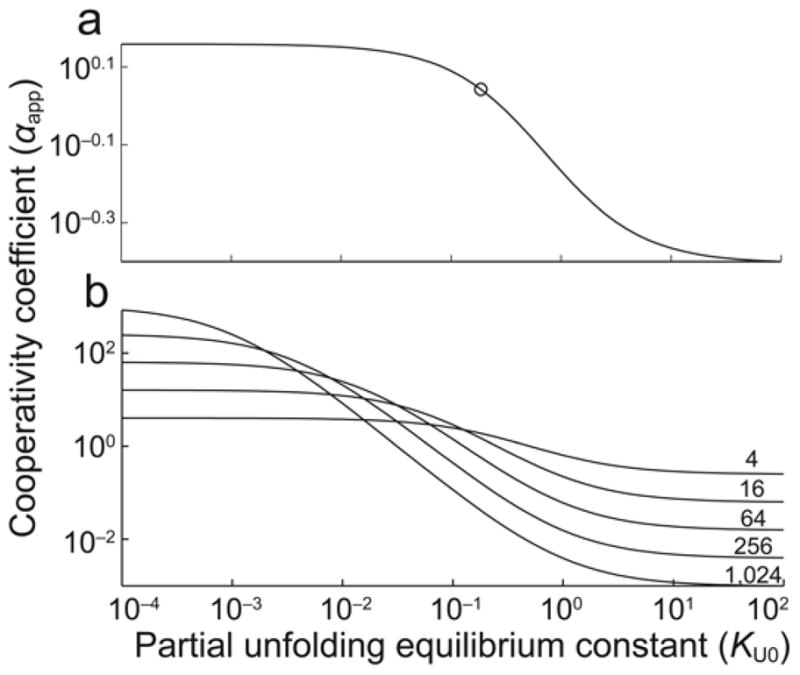
Allostery has been studied for many decades, yet it remains challenging to determine experimentally how it occurs at a molecular level. We have developed an approach combining isothermal titration calorimetry, circular dichroism and nuclear magnetic resonance spectroscopy to quantify allostery in terms of protein thermodynamics, structure and dynamics. This strategy was applied to study the interaction between aminoglycoside N-(6')-acetyltransferase-Ii and one of its substrates, acetyl coenzyme A. It was found that homotropic allostery between the two active sites of the homodimeric enzyme is modulated by opposing mechanisms. One follows a classical Koshland-Némethy-Filmer (KNF) paradigm, whereas the other follows a recently proposed mechanism in which partial unfolding of the subunits is coupled to ligand binding. Competition between folding, binding and conformational changes represents a new way to govern energetic communication between binding sites.
Figures
References
-
- Perutz MF. Mechansism of cooperativity and allosteric regulation in proteins. Q Rev Biophys. 1989;22:139–236. - PubMed
-
- Monod J, Wyman J, Changeux JP. On nature of allosteric transitions - a plausible model. J Mol Biol. 1965;12:88–118. - PubMed
-
- DeDecker BS. Allosteric drugs: thinking outside the active-site box. Chem Biol. 2000;7:R103–R107. - PubMed
-
- Koshland DE, Nemethy G, Filmer D. Comparison of experimental binding data and theoretical models in proteins containing subunits. Biochemistry. 1966;5:365–385. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
