

Comparative analysis of carboxysome shell proteins
- PMID: 21279737
- PMCID: PMC3173617
- DOI: 10.1007/s11120-011-9624-6
Comparative analysis of carboxysome shell proteins
Abstract
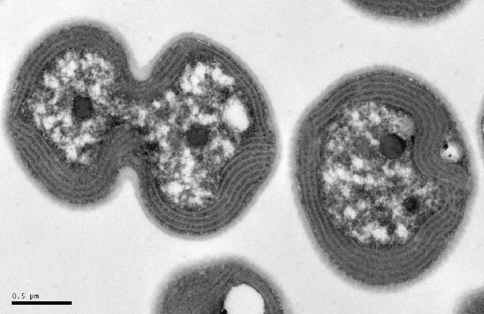
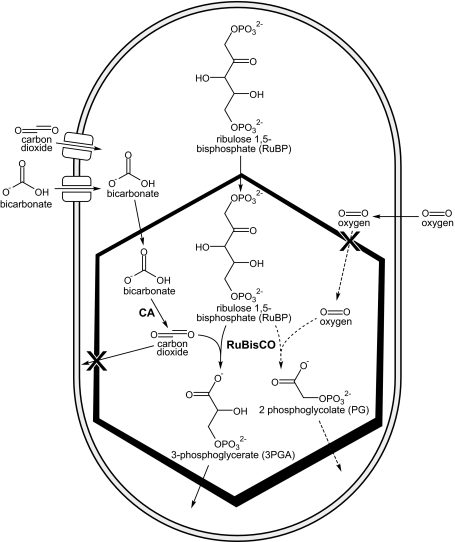
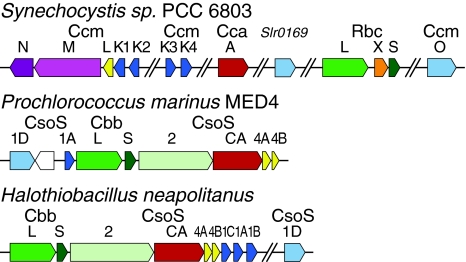
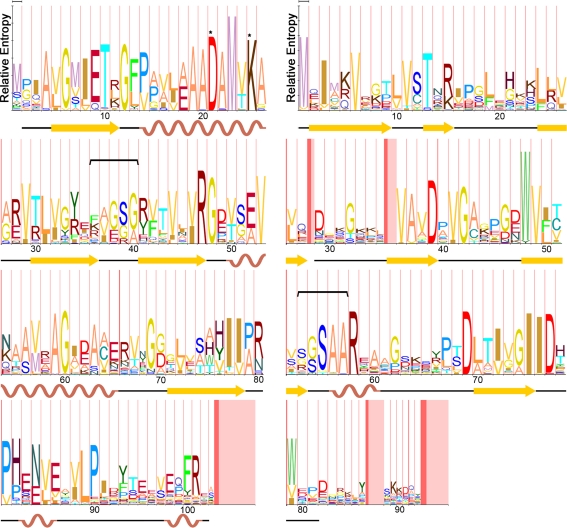
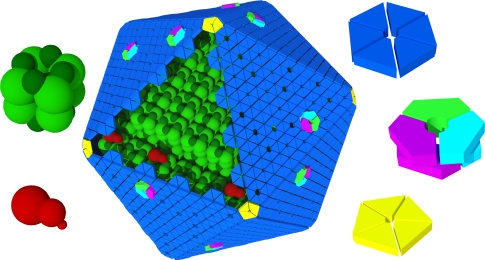
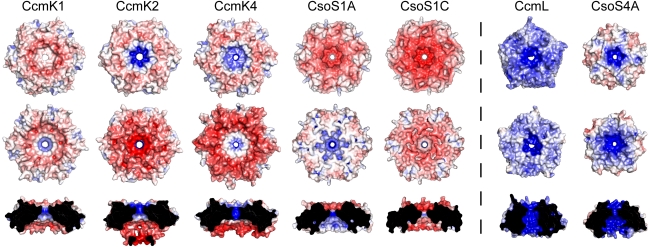
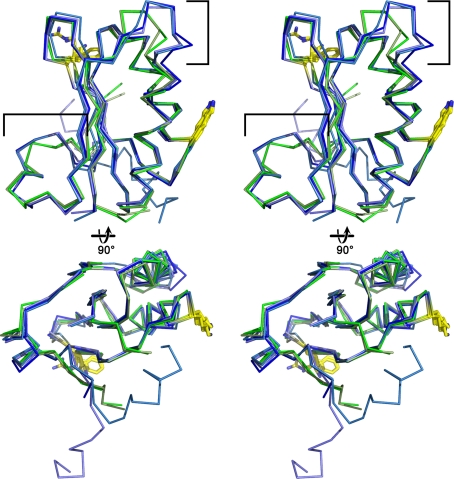
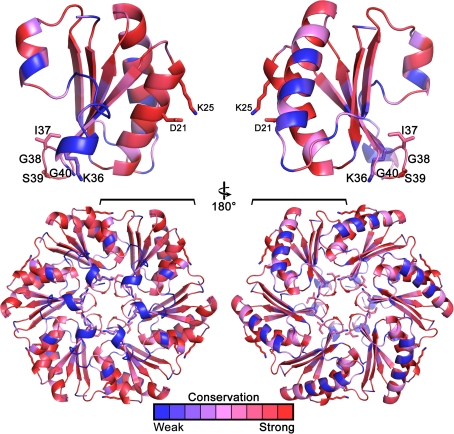
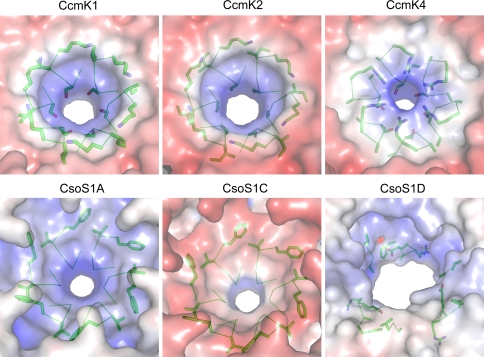
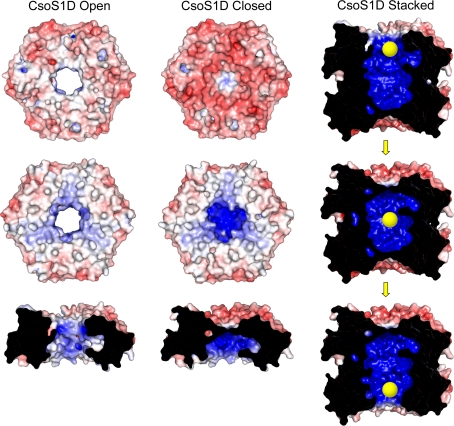
Carboxysomes are metabolic modules for CO(2) fixation that are found in all cyanobacteria and some chemoautotrophic bacteria. They comprise a semi-permeable proteinaceous shell that encapsulates ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) and carbonic anhydrase. Structural studies are revealing the integral role of the shell protein paralogs to carboxysome form and function. The shell proteins are composed of two domain classes: those with the bacterial microcompartment (BMC; Pfam00936) domain, which oligomerize to form (pseudo)hexamers, and those with the CcmL/EutN (Pfam03319) domain which form pentamers in carboxysomes. These two shell protein types are proposed to be the basis for the carboxysome's icosahedral geometry. The shell proteins are also thought to allow the flux of metabolites across the shell through the presence of the small pore formed by their hexameric/pentameric symmetry axes. In this review, we describe bioinformatic and structural analyses that highlight the important primary, tertiary, and quaternary structural features of these conserved shell subunits. In the future, further understanding of these molecular building blocks may provide the basis for enhancing CO(2) fixation in other organisms or creating novel biological nanostructures.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
