

Expression, secretion and surface display of a human alkaline phosphatase by the ciliate Tetrahymena thermophila
- PMID: 21281462
- PMCID: PMC3042934
- DOI: 10.1186/1472-6750-11-11
Expression, secretion and surface display of a human alkaline phosphatase by the ciliate Tetrahymena thermophila
Abstract
Background: Tetrahymena thermophila possesses many attributes that render it an attractive host for the expression of recombinant proteins. Surface proteins from the parasites Ichthyophthirius multifiliis and Plasmodium falciparum and avian influenza virus antigen H5N1 were displayed on the cell membrane of this ciliate. Furthermore, it has been demonstrated that T. thermophila is also able to produce a functional human DNase I. The present study investigates the heterologous expression of the functional human intestinal alkaline phosphatase (hiAP) using T. thermophila and thereby presents a powerful tool for the optimization of the ciliate-based expression system.
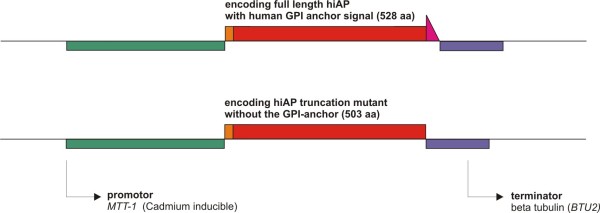
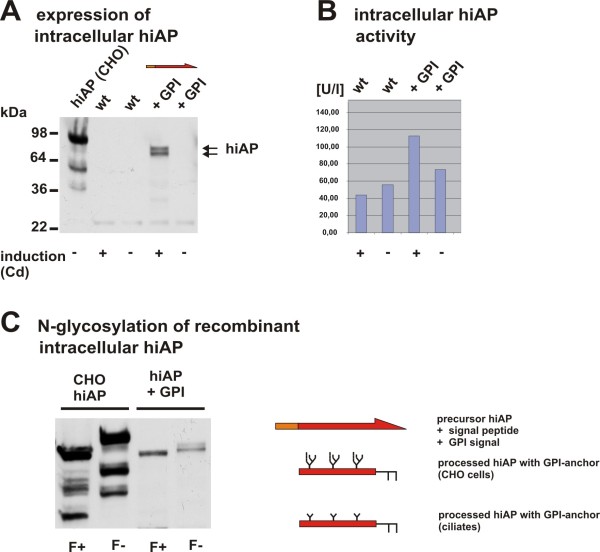
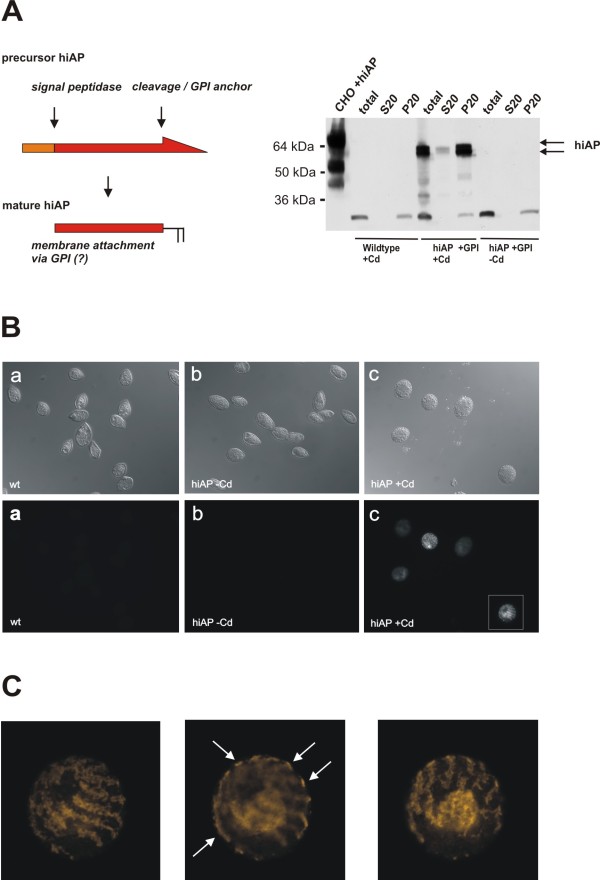
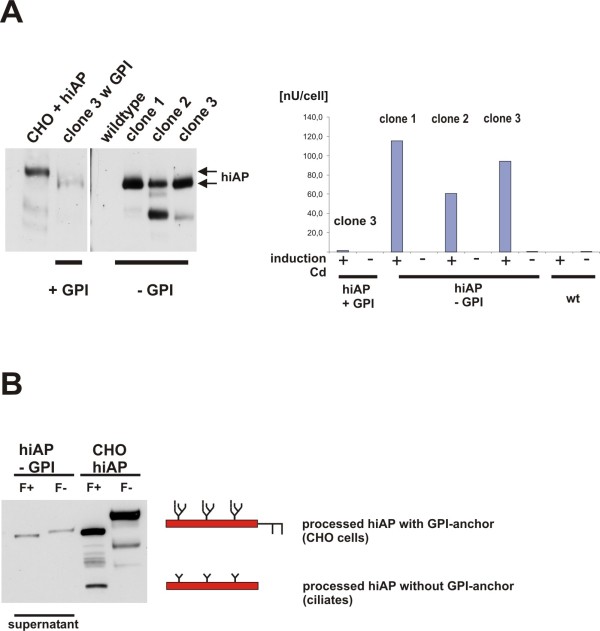
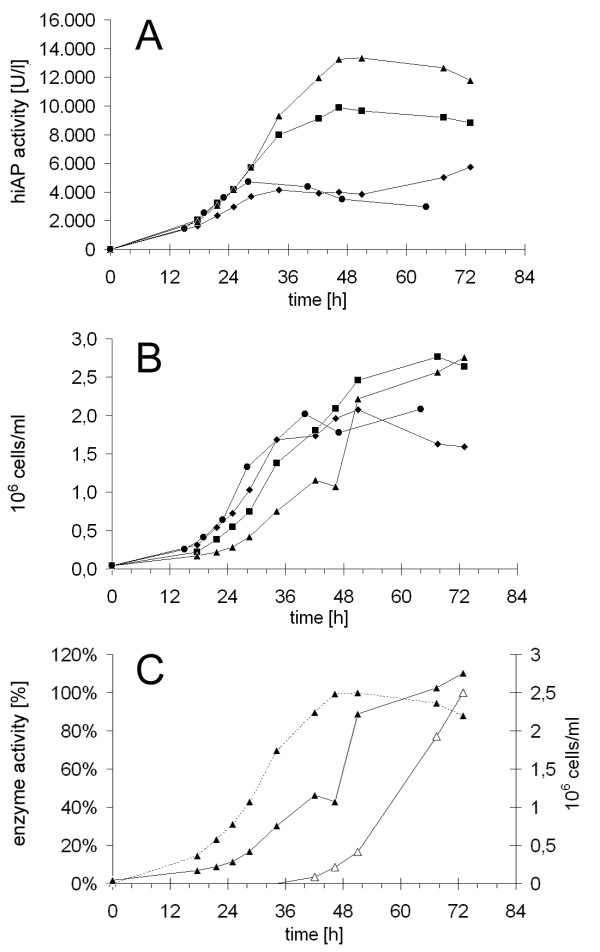
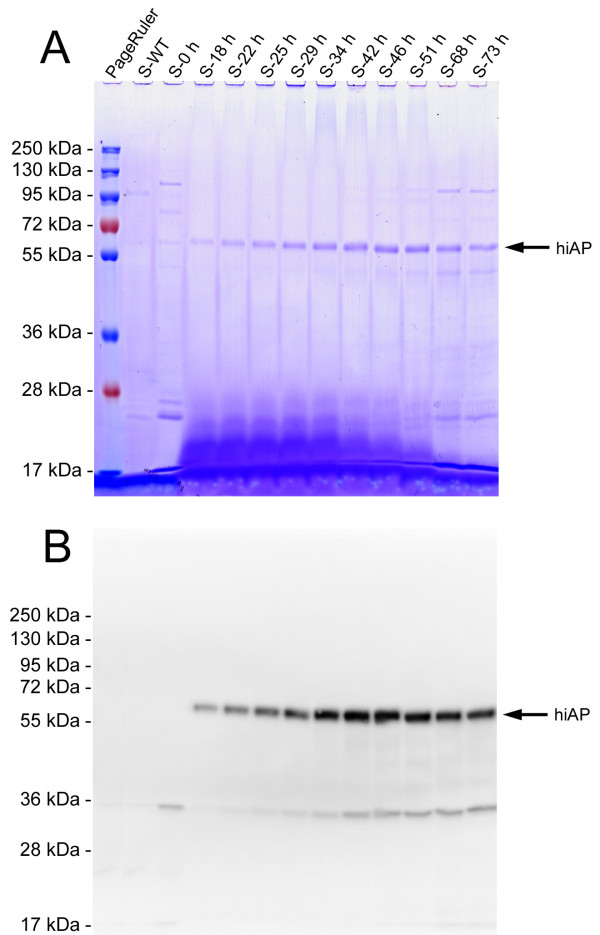
Results: Functional and full length human intestinal alkaline phosphatase was expressed by T. thermophila using a codon-adapted gene containing the native signal-peptide and GPI (Glycosylphosphatidylinositol) anchor attachment signal. HiAP activity in the cell extract of transformants suggested that the hiAP gene was successfully expressed. Furthermore, it was demonstrated that the enzyme was modified with N-glycosylation and localized on the surface membrane by the C-terminal GPI anchor. A C-terminally truncated version of hiAP lacking the GPI anchor signal peptide was secreted into the medium as an active enzyme. In a first approach to establish a high level expression system up to 14,000 U/liter were produced in a time frame of two days, which exceeds the production rate of other published expression systems for this enzyme.
Conclusions: With the expression of hiAP, not only a protein of commercial interest could be produced, but also a reporter enzyme that offers the possibility to analyze T. thermophila genes that play a role in the regulation of protein secretion. Additionally, the fact that ciliates do not secrete an endogenous alkaline phosphatase provides the possibility to use the truncated hiAP as a reporter enzyme, allowing the quantification of measures that will be necessary for further optimization of the host strains and the fermentation processes.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
