

Accelerated human mutant tau aggregation by knocking out murine tau in a transgenic mouse model
- PMID: 21281813
- PMCID: PMC3069830
- DOI: 10.1016/j.ajpath.2010.10.034
Accelerated human mutant tau aggregation by knocking out murine tau in a transgenic mouse model
Abstract
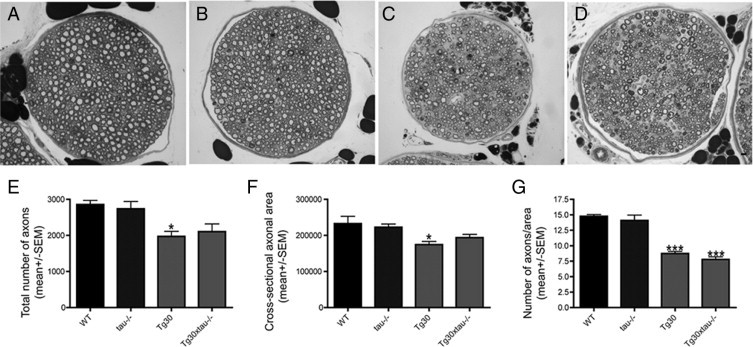
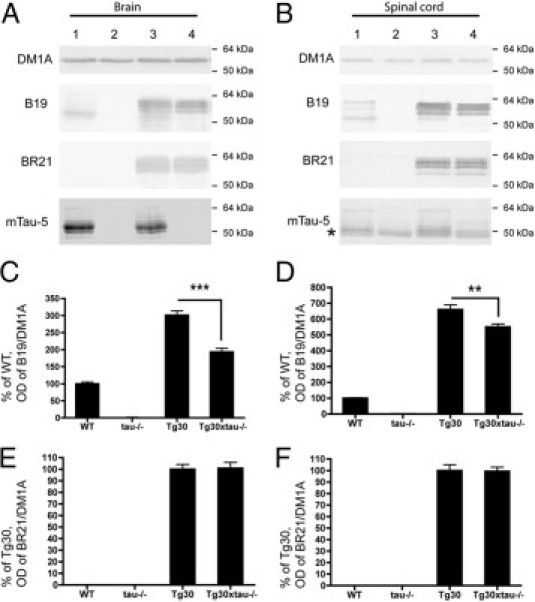
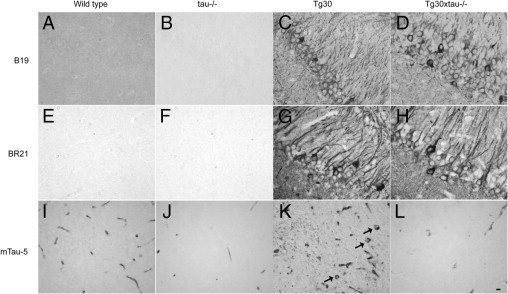
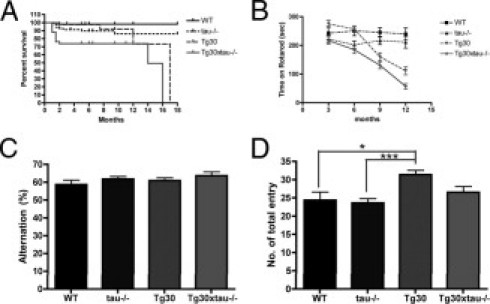
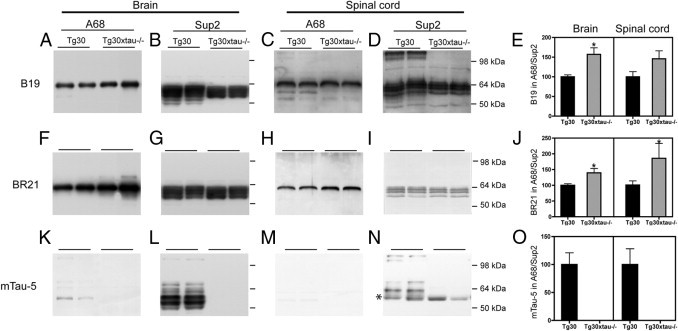
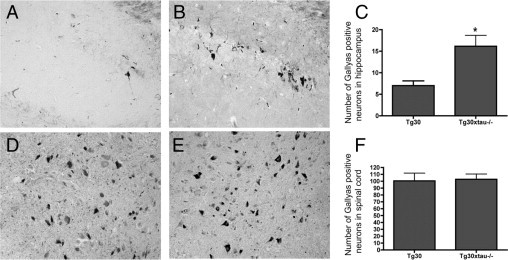
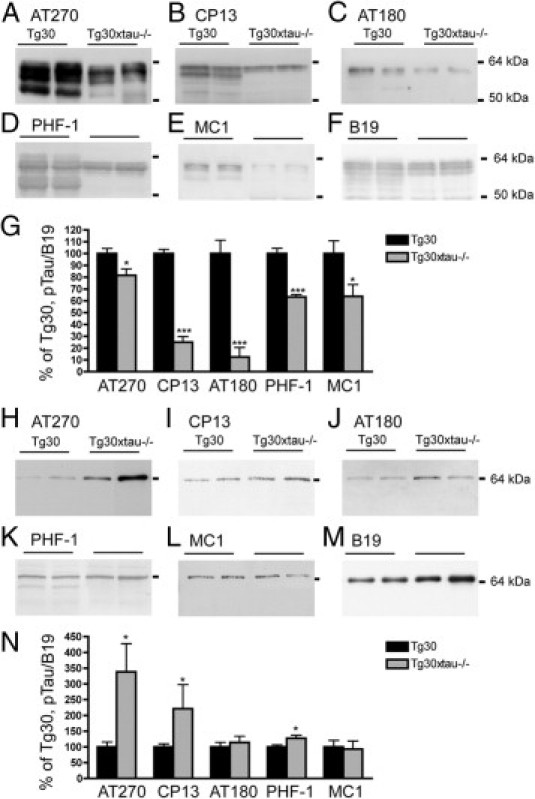
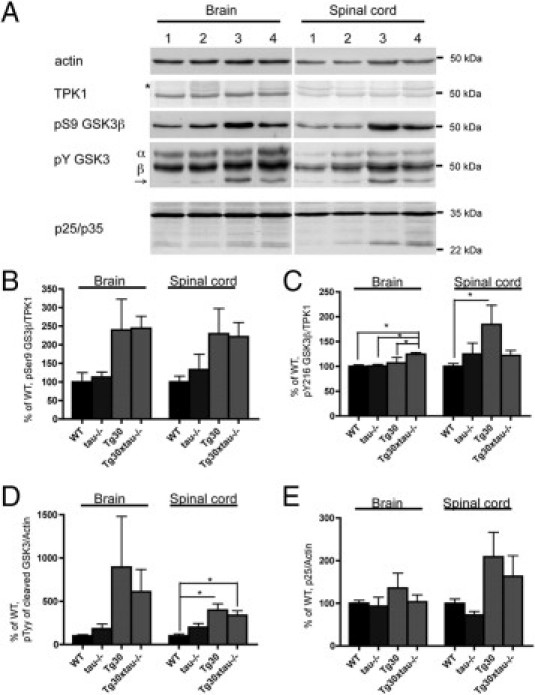
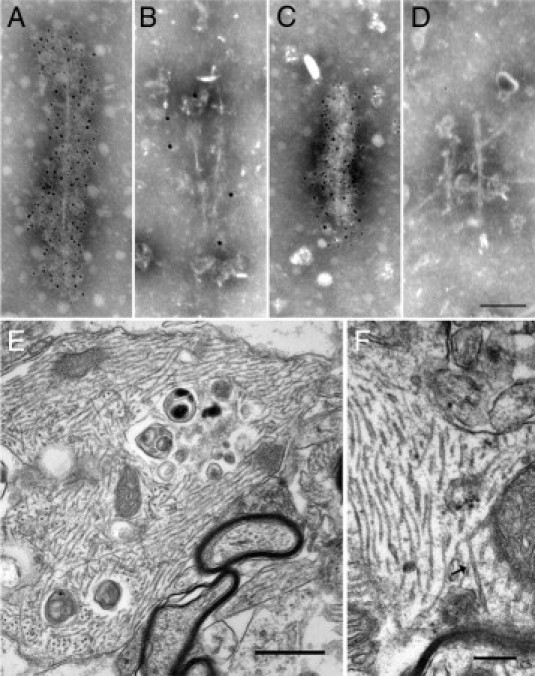
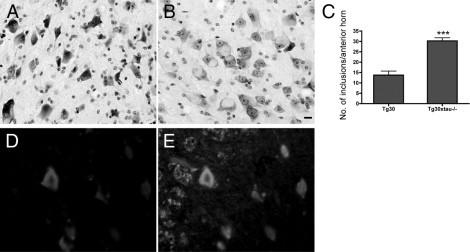
Many models of human tauopathies have been generated in mice by expression of a human mutant tau with maintained expression of mouse endogenous tau. Because murine tau might interfere with the toxic effects of human mutant tau, we generated a model in which a pathogenic human tau protein is expressed in the absence of wild-type tau protein, with the aim of facilitating the study of the pathogenic role of the mutant tau and to reproduce more faithfully a human tauopathy. The Tg30 line is a tau transgenic mouse model overexpressing human 1N4R double-mutant tau (P301S and G272V) that develops Alzheimer's disease-like neurofibrillary tangles in an age-dependent manner. By crossing Tg30 mice with mice invalidated for their endogenous tau gene, we obtained Tg30xtau(-/-) mice that express only exogenous human double-mutant 1N4R tau. Although Tg30xtau(-/-) mice express less tau protein compared with Tg30, they exhibit signs of decreased survival, increased proportion of sarkosyl-insoluble tau in the brain and in the spinal cord, increased number of Gallyas-positive neurofibrillary tangles in the hippocampus, increased number of inclusions in the spinal cord, and a more severe motor phenotype. Deletion of murine tau accelerated tau aggregation during aging of this mutant tau transgenic model, suggesting that murine tau could interfere with the development of tau pathology in transgenic models of human tauopathies.
Copyright © 2011 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Selkoe D.J. The origins of Alzheimer disease—A is for amyloid. JAMA. 2000;283:1615–1617. - PubMed
-
- Ballatore C., Lee V.M.Y., Trojanowski J.Q. Tau-mediated neurodegeneration in Alzheimer's disease and related disorders. Nat Rev Neurosci. 2007;8:663–672. - PubMed
-
- Lewis J., Dickson D.W., Lin W.L., Chisholm L., Corral A., Jones G., Yen S.H., Sahara N., Skipper L., Yager D., Eckman C., Hardy J., Hutton M., McGowan E. Enhanced neurofibrillary degeneration in transgenic mice expressing mutant tau and APP. Science. 2001;293:1487–1491. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
