

The role of synaptobrevin1/VAMP1 in Ca2+-triggered neurotransmitter release at the mouse neuromuscular junction
- PMID: 21282288
- PMCID: PMC3099018
- DOI: 10.1113/jphysiol.2010.201939
The role of synaptobrevin1/VAMP1 in Ca2+-triggered neurotransmitter release at the mouse neuromuscular junction
Abstract
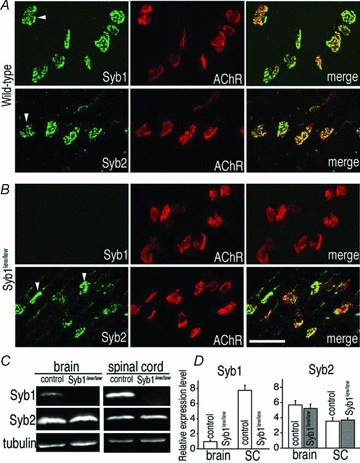
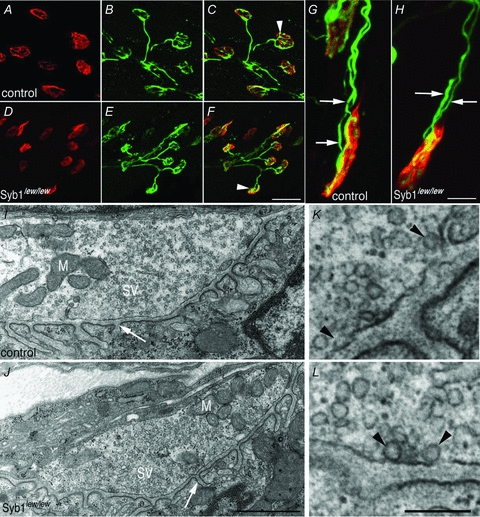
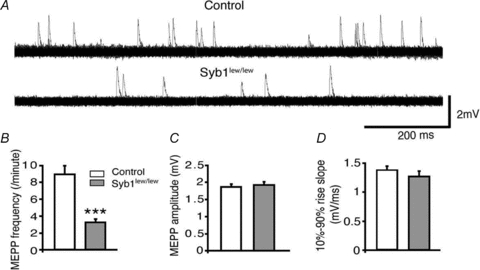
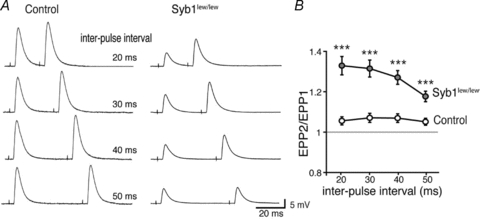
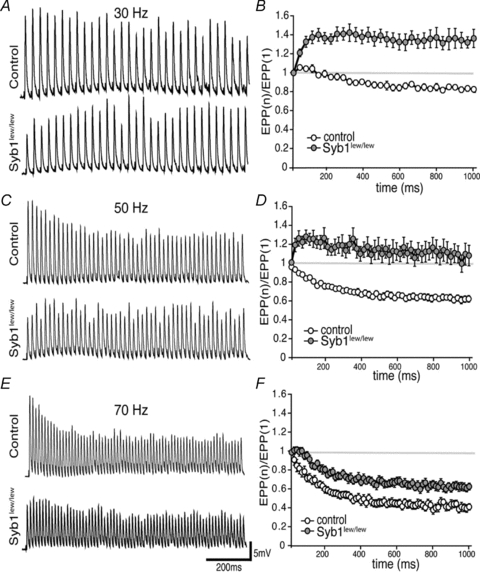
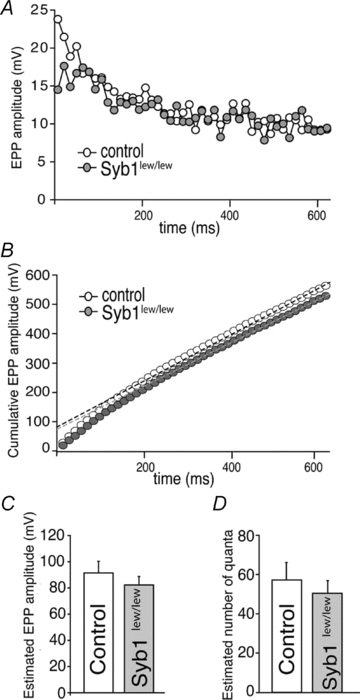
Synaptobrevin (Syb)/vesicle-associated membrane protein (VAMP) is a small, integral membrane protein of synaptic vesicles. Two homologous isoforms of synaptobrevin, Syb1/VAMP1 and Syb2/VAMP2, exhibit distinct but partially overlapping patterns of expression in adult mammalian neurons: Syb1 is predominantly expressed in the spinal cord, especially in motor neurons and motor nerve terminals of the neuromuscular junction (NMJ), whereas Syb2 is primarily expressed in central synapses in the brain. Whereas many studies have focused on the function of Syb2 in the brain, few studies have examined the role of Syb1. Here we report that Syb1 plays a critical role in neuromuscular synaptic transmission. A null mutation of Syb1 resulting from a spontaneous, nonsense mutation in mice significantly impairs the function, but not the structure, of the NMJ. In particular, both spontaneous and evoked synaptic activities in Syb1 mutant mice are reduced significantly relative to control mice. Short-term synaptic plasticity in Syb1-deficient NMJs is markedly altered: paired-pulse facilitation is significantly enhanced, suggesting a reduction in the initial release probability of synaptic vesicles. Furthermore, Syb1-deficient NMJs display a pronounced asynchrony in neurotransmitter release. These impairments are not due to an alteration of the size of the readily releasable pool of vesicles, but are attributable to reduced sensitivity and cooperativity to calcium (Ca2+) due to the absence of Syb1. Our findings demonstrate that Syb1 plays an essential, non-redundant role in Ca2+-triggered vesicle exocytosis at the mouse NMJ.
Figures
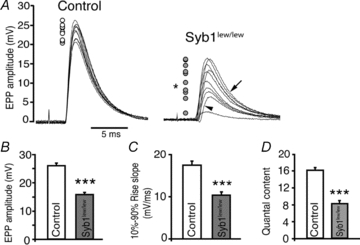
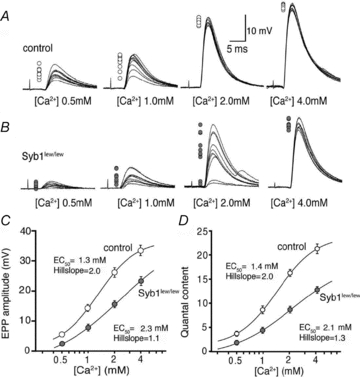
 , where Y is the value of the EPP amplitude or the quantal content at the indicated [Ca2+], Ymax and Ymin are the maximum and minimum EPP amplitude or quantal content, respectively, EC50 is the [Ca2+] when Y is 50% of Ymax, and k is the Hill coefficient (Hill slope), which reflects the steepness of the curve. The sigmoidal dose–response curve of EPP amplitude and quantal content as a function of external [Ca2+] showed that the EC50 was markedly increased and the Hill slope was markedly reduced in Syb1lew/lew NMJs (n = 29) compared with the control NMJs (n = 23).
, where Y is the value of the EPP amplitude or the quantal content at the indicated [Ca2+], Ymax and Ymin are the maximum and minimum EPP amplitude or quantal content, respectively, EC50 is the [Ca2+] when Y is 50% of Ymax, and k is the Hill coefficient (Hill slope), which reflects the steepness of the curve. The sigmoidal dose–response curve of EPP amplitude and quantal content as a function of external [Ca2+] showed that the EC50 was markedly increased and the Hill slope was markedly reduced in Syb1lew/lew NMJs (n = 29) compared with the control NMJs (n = 23).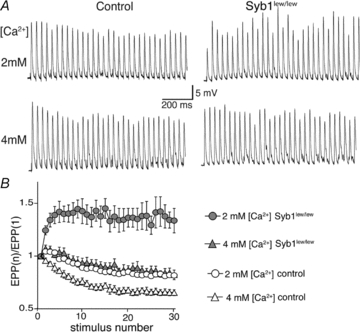
References
-
- Arac D, Chen X, Khant HA, Ubach J, Ludtke SJ, Kikkawa M, Johnson AE, Chiu W, Sudhof TC, Rizo J. Close membrane-membrane proximity induced by Ca2+-dependent multivalent binding of synaptotagmin-1 to phospholipids. Nat Struct Mol Biol. 2006;13:209–217. - PubMed
-
- Augustine GJ, Charlton MP, Smith SJ. Calcium action in synaptic transmitter release. Annu Rev Neurosci. 1987;10:633–693. - PubMed
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous