

Metabolic impact of adult-onset, isolated, growth hormone deficiency (AOiGHD) due to destruction of pituitary somatotropes
- PMID: 21283519
- PMCID: PMC3023710
- DOI: 10.1371/journal.pone.0015767
Metabolic impact of adult-onset, isolated, growth hormone deficiency (AOiGHD) due to destruction of pituitary somatotropes
Abstract
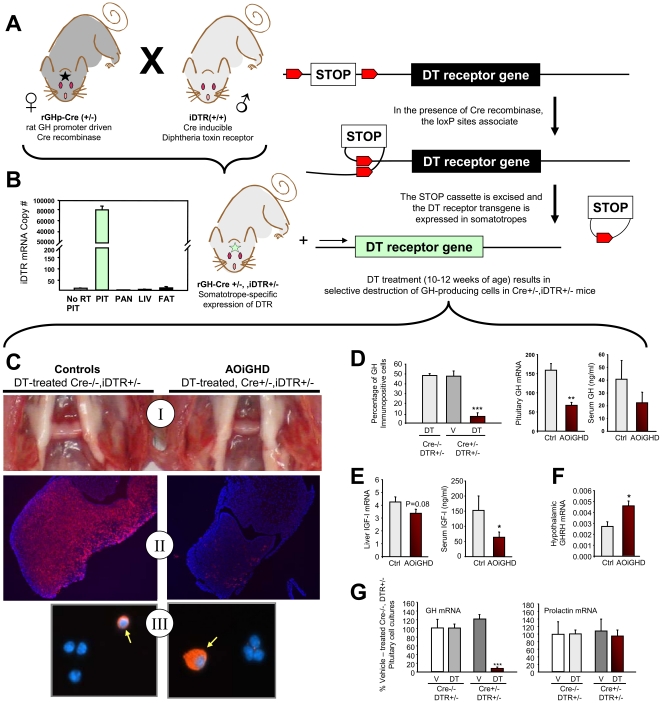
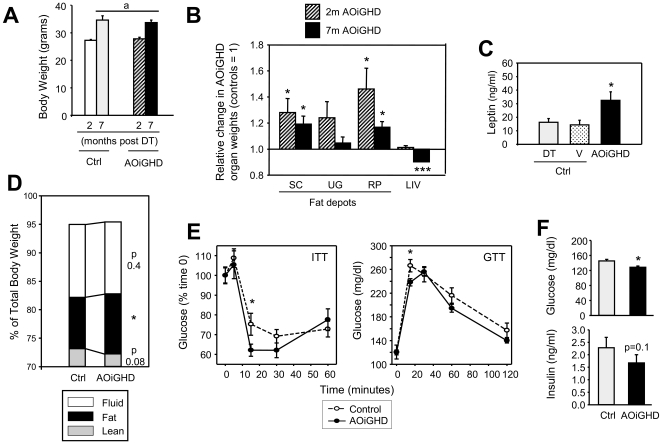
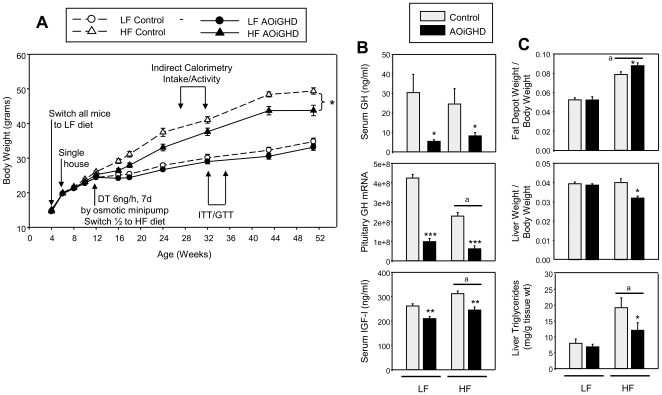
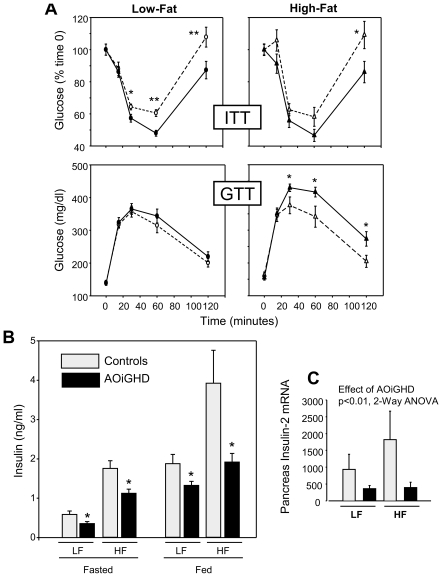
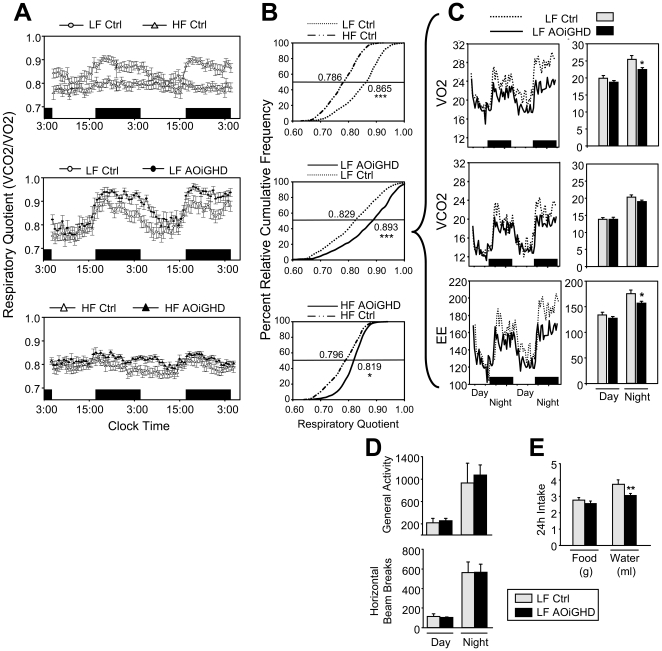
Growth hormone (GH) inhibits fat accumulation and promotes protein accretion, therefore the fall in GH observed with weight gain and normal aging may contribute to metabolic dysfunction. To directly test this hypothesis a novel mouse model of adult onset-isolated GH deficiency (AOiGHD) was generated by cross breeding rat GH promoter-driven Cre recombinase mice (Cre) with inducible diphtheria toxin receptor mice (iDTR) and treating adult Cre(+/-),iDTR(+/-) offspring with DT to selectively destroy the somatotrope population of the anterior pituitary gland, leading to a reduction in circulating GH and IGF-I levels. DT-treated Cre(-/-),iDTR(+/-) mice were used as GH-intact controls. AOiGHD improved whole body insulin sensitivity in both low-fat and high-fat fed mice. Consistent with improved insulin sensitivity, indirect calorimetry revealed AOiGHD mice preferentially utilized carbohydrates for energy metabolism, as compared to GH-intact controls. In high-fat, but not low-fat fed AOiGHD mice, fat mass increased, hepatic lipids decreased and glucose clearance and insulin output were impaired. These results suggest the age-related decline in GH helps to preserve systemic insulin sensitivity, and in the context of moderate caloric intake, prevents the deterioration in metabolic function. However, in the context of excess caloric intake, low GH leads to impaired insulin output, and thereby could contribute to the development of diabetes.
Conflict of interest statement
Figures
References
-
- Giustina A, Veldhuis JD. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocr Rev. 1998;19:717–797. - PubMed
-
- Scacchi M, Pincelli AI, Cavagnini F. Growth hormone in obesity. Int J Obes Relat Metab Disord. 1999;23:260–271. - PubMed
-
- Attallah H, Friedlander AL, Hoffman AR. Visceral obesity, impaired glucose tolerance, metabolic syndrome, and growth hormone therapy. Growth Horm IGF Res. 2006;16:62–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
